氮、磷添加对不同种植密度樟树幼苗碳储量及其分配的影响
2020-01-18余明蔡金桓薛立
余明,蔡金桓,薛立
(华南农业大学林学与风景园林学院,广东广州510642)
氮(N)和磷(P)是影响植物生命活动的必要元素,对调节植物生长发育有重要意义[1]。热带和亚热带地区的土壤风化程度较高导致了土壤磷的缺乏,引起该地区的陆地生态系统受到磷的限制[2]。我国华南地区位于亚热带,近年来的社会发展和人类活动导致了氮沉降的持续发生[3],可能会进一步加剧磷的限制作用。华南地区林业生产上,经常通过施加磷肥缓解森林土壤的磷缺乏,因此氮、磷添加对于林木生长发育的影响受到广泛关注。
碳(C)是组成植物体的重要结构性物质[4],在调节林木生长发育和维持生态系统平衡方面发挥着关键作用[5-6]。研究表明,氮和磷等营养元素的输入能够影响植物的碳累积[7],适度的氮沉降通常有利于植物的生长发育,而长期过量的氮沉降则抑制植物生长,引起土壤酸化并减弱生态系统的固碳能力[8]。磷添加可以缓解生态系统的磷限制,并提高生态系统的生产力。当前,磷沉降的研究主要集中在水生态系统[9],鲜有森林生态系统磷沉降的报道。近年来,国内对森林生态系统碳储量的研究主要集中在不同林龄[10]、林分类型[11]和土壤[12]等方面,氮沉降对于森林碳储量的影响仍存在争议[13-14],鲜有磷添加影响森林碳储量的报道。
密度是自然界中重要的选择压力,林分密度影响了植物对光照和养分的吸收利用,并引起相邻植物间对资源的激烈竞争,从而改变森林生物量的积累与分配[15]。当前,关于密度对植物幼苗的影响已有少量研究[16-17],但鲜见氮、磷添加与密度的交互作用对幼苗碳储量影响的报道。樟树Cinnamomum camphora是我国特有的珍贵用材和经济树种[18],前人主要研究了樟树的光合作用[19]、生理[20]、土壤碳氮储量[21]、施氮对土壤氮矿化的影响[22]和凋落物量及养分动态特征等[23]。本研究以1年生樟树幼苗为试验材料,通过分析氮、磷添加和不同种植密度对樟树幼苗各器官碳储量和分配格局的影响,以期了解氮磷沉降和密度影响樟树碳分配的机制,为氮磷沉降环境下亚热带地区樟树林的可持续经营提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于广东省广州市华南农业大学试验林(113°21′E,23°09′N),该地年平均气温21.9℃,平均相对湿度77%,年降雨量约为1 736mm,全年水热同期,雨量充沛,属亚热带季风气候。试验地光照充足,适合幼苗的生长,土壤类型为砂页岩发育的酸性红壤,土壤磷含量较低。处理前土壤的pH 为4.02,有机质、全氮和全磷的质量分数分别为14.27、0.62和0.17 g·kg−1,碱解氮和有效磷的质量分数分别为57.40和0.10mg·kg−1。
1.2 试验设计
以广东国森林业有限公司所提供的樟树1年生实生苗为试验材料,幼苗平均株高0.47m,平均地径0.36 cm,平均冠幅11.10 cm。采用直径35 cm,深30 cm 的无纺布美植袋种植,基质为该试验地0~20 cm 土层的土壤。本试验选择氯化铵(NH4Cl)模拟大气氮沉降,以二水合磷酸二氢钠(NaH2PO4·2H2O)进行磷添加。根据样地的氮沉降水平背景值以及参考同类研究方法[24-25],并考虑到未来的氮沉降持续增加,确定N和P的添加量。
试验时间为2017年6—9月。试验采用4×4双因素析因设计。N、P添加处理设置4个水平:不加N和P(对照,CK),加N,加P,加N+P。N、P及N+P每年的添加量分别为NH4C l 40 g·m−2,NaH2PO4·2H2O 20 g·m−2,NH4C l 40 g·m−2+NaH2PO4·2H2O 20 g·m−2。种植密度设置4个水平:10、20、40和80株·m−2。试验采用完全随机区组设计,共16个处理,每处理3次重复,共48个小区,每个小区16株幼苗。根据处理水平的要求,N、P添加时,将各处理每次所需质量的肥料溶于12 L水中,向每袋的幼苗浇灌200mL 溶液,保证不会引起烧苗的现象,不施肥处理的只浇灌相同量的水。自6月起的每月月初和月中分2次向幼苗施肥直至收获,共施肥8次。幼苗处理前的平均苗高和平均地径分别为(0.47±0.03)和(0.36±0.07)cm。处理前的幼苗各器官生物量和碳含量见表1。

表1 幼苗处理前各器官单株生物量和碳含量(平均值±标准误差)Table 1 Biomassand carbon content per seed ling before treatment (mean ± SE)
1.3 研究方法
2017年9月底,取16种处理的幼苗各15株,将植株分解为根、茎、枝和叶4部分,称量各器官鲜质量,再将其分别装入信封中编号,置烘箱中105℃条件下杀青2 h,转至80℃烘24 h 至恒质量,称量各器官干质量,求出单株幼苗各器官的生物量,再根据种植密度估算出单位面积各器官的生物量。将不同处理幼苗的根、茎、枝和叶样品充分粉碎后,过60目筛,单株各器官分别取10 g 样品,用重铬酸钾外加热法测定碳含量[26-27],计算幼苗各器官碳储量(碳储量=生物量×碳含量)。土壤pH 用电位法测定,用重铬酸钾氧化−外加热法测定土壤有机质含量,土壤全N 和全P分别用半微量凯氏法和钼蓝比色法测定,土壤碱解N 和有效P含量分别采用碱解−扩散法和0.5mol·L−1碳酸氢钠法测定[26]。
1.4 数据处理
用Microsoft Excel2003对数据进行平均值、标准误差分析和作图。用SAS 9.3统计分析软件对植物各器官碳含量和碳储量进行方差分析,而后对有显著差异的数据进行Duncan’s多重检验,P<0.05为差异显著。
2 结果与分析
2.1 氮、磷添加对不同种植密度樟树幼苗各器官碳含量的影响
试验结束时,各添加处理下的幼苗茎、枝、根的C含量差异不大。在各种植密度条件下,N 和N+P添加处理下的幼苗叶片C含量均高于不加N和P处理(CK)。此外,随着种植密度的增大,叶片C含量表现出下降的趋势(表2)。

表2 不同种植密度及氮、磷添加处理幼苗各器官的碳含量1)(平均值±标准误差)Table 2 Carbon contentsof organs per seed ling under different p lanting densitiesand nitrogen and phosphorusadditions(mean±SE)
2.2 氮、磷添加对不同种植密度樟树幼苗各器官碳储量的影响
与对照相比,N、P添加处理对单株幼苗C总储量整体表现为促进作用,但N、P添加各处理间差异不显著。随着种植密度的增大,相同N、P添加处理的单株幼苗C总储量呈现下降的趋势(表3)。

表3 不同种植密度及氮、磷添加处理下单株幼苗各器官的碳储量和分配比例1)Table 3 Carbon storage of organs and its allocation per seed ling under different p lanting densities and nitrogen and phosphorus additions
各处理的单株幼苗各器官的C储量分配表现各异。CK 处理各密度单株幼苗的C储量的分配比例呈现为根>茎>叶>枝;对于N 添加处理,在种植密度10和20株·m−2条件下,单株幼苗C储量分配比例呈现为根>叶>茎>枝,种植密度40株·m−2条件下其呈现为根>茎>叶>枝,种植密度80株·m−2条件下其呈现为根>茎>枝>叶;对于P添加处理,在种植密度10株·m−2条件下,单株幼苗C储量分配比例呈现为叶>根>茎>枝,种植密度20和40株·m−2条件下其呈现为根>茎>叶>枝,种植密度80株·m−2条件下其呈现为茎>根>枝>叶;对于N+P添加处理,在种植密度10和20株·m−2条件下,单株幼苗C储量的分配比例呈现为叶>根>茎>枝,种植密度40和80株·m−2条件下的单株幼苗C储量分配比例呈现为根>茎>叶>枝。可见,P和N+P添加处理在低种植密度条件下,单株幼苗叶的C储量分配比例相对较高,高密度种植则促进了C储量向茎进行分配。
由表4的方差分析可知,单株幼苗各器官C储量和总C储量在各种植密度间和不同N、P添加处理间均存在显著性差异,种植密度对单株幼苗及各器官C储量的影响均大于各N、P添加处理。种植密度和N、P添加处理的交互作用对单株幼苗的枝、叶和总C 储量有显著影响。
2.3 氮、磷添加和不同种植密度处理下樟树幼苗各器官的单位面积碳储量
如表5所示,各N、P添加处理下幼苗各器官的单位面积C储量有所差异。总体而言,N、P添加处理能够促进幼苗的单位面积C储量增加,但是随着种植密度的增大,其促进效果逐渐减弱。

表4 不同种植密度和氮、磷添加对单株幼苗各器官碳储量交互作用的方差分析Table 4 Variance analyses of interaction effects of planting density and nitrogen and phosphorus additions on carbon storage of organsper seed ling

表5 不同种植密度及氮、磷添加处理下幼苗各器官的单位面积碳储量和分配比例1)Table 5 Carbon storage of organs and its allocation in unit area under different p lanting densities and nitrogen and phosphorus additions
C K 处理情况下,种植密度1 0、2 0和4 0株·m−2时幼苗各器官的单位面积C储量呈现为根>茎>叶>枝,种植密度80株·m−2时其呈现为根>茎>枝>叶;N 添加处理情况下,种植密度10和20株·m−2时幼苗各器官单位面积C储量为根>叶>茎>枝,种植密度40株·m−2时其呈现为根>茎>叶>枝,种植密度80株·m−2时其呈现为根>茎>枝>叶;P添加处理情况下,种植密度10株·m−2时幼苗各器官单位面积C 储量呈现为叶>根>茎>枝,种植密度20和40株·m−2时其呈现为根>茎>叶>枝,种植密度80株·m−2时其呈现为茎>根>枝>叶;N+P添加处理下,种植密度10、20株·m−2时幼苗各器官单位面积C储量为叶>根>茎>枝,种植密度40和80株·m−2时其呈现为根>茎>叶>枝。总体而言,在各种植密度条件下,氮、磷添加处理的幼苗各器官的单位面积C 储量基本上均有所增加。氮、磷添加对低种植密度下幼苗叶的单位面积C储量分配比例有促进效果,在高种植密度条件下,幼苗叶的单位面积C储量分配比例降低。
由表6可知,单株幼苗及根、茎和枝的单位面积C储量在各种植密度间和不同N、P添加处理间均有显著性差异,幼苗叶的单位面积C储量只在各氮、磷添加处理间有显著性差异,其中种植密度对单株幼苗及根茎和枝的单位面积C储量的作用较强,N、P添加处理对幼苗叶有较大影响。种植密度和N、P添加处理的交互作用对幼苗枝和叶的单位面积C储量有显著影响。
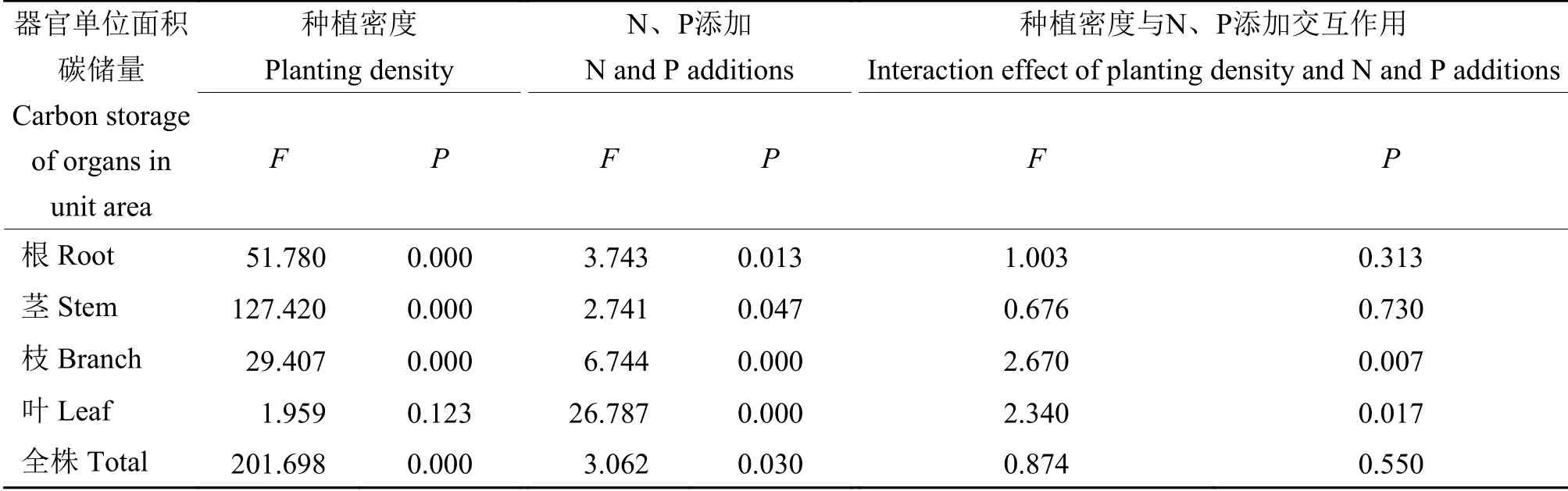
表6 不同种植密度及氮、磷添加对幼苗各器官的单位面积碳储量交互作用的方差分析Table 6 Variance analyses of interaction effects of planting density and nitrogen and phosphorus additions on carbon storage of organs in unit area
3 讨论与结论
3.1 氮、磷添加对于不同种植密度樟树幼苗碳含量与分配的影响
本研究结果显示,N、P添加处理对幼苗根、茎和枝的C含量影响不显著,这与Currey 等[28]的研究结果相类似。原因可能是植物体内的功能性物质含量受环境影响较大,而结构性物质受影响较小的缘故。C 是组成植物体的结构性物质,故其含量较为稳定。此外,C通常不是植物生长受限的主要元素,所以其在大部分植物体内的含量较高且变化较小[29]。翁俊等[30]的研究结果表明,N 沉降能促进毛竹Phyllostachysedulis叶的C含量增加,本研究也发现N 和N+P添加处理在低密度种植时能够促使樟树幼苗叶的C含量上升。因为叶片是光合固碳的重要器官,N 添加对植物叶片N 素、叶绿素和光合色素含量[31]以及Rubisco酶浓度[32]有促进作用,从而提高植物的光合速率,最终引起叶C含量的升高。另外,N 和P的输入可以缓解土壤的养分限制,从而增强植物根系对营养元素的吸收能力[33],间接促进植物叶片光合作用的固C能力。
密度影响着植物个体生长过程中可获取的资源环境空间[34]。通常情况下,种植密度越大,资源对植物的限制作用越大[15]。本研究发现随着种植密度的增大,樟树幼苗叶片C含量表现出下降的趋势。这是因为高种植密度下个体间对养分的争夺较为激烈,并且植株间距小导致叶片较多地重叠和相互遮蔽,从而引起植物光合速率下降,造成植物叶片光合固碳能力的降低[35]。
3.2 氮、磷添加对不同种植密度樟树幼苗碳储量与分配的影响
本研究表明,相比于对照,N、P添加处理能够促进幼苗平均单株和单位面积C总储量增加,这与Yan 等[7]的研究结果相类似,原因可能是N的输入提升了植物的净初级生产力[36],从而提高幼苗的固C量。植物体内蛋白质的合成需要光合作用同化的C作为主要组分,而适当的P输入有助于蛋白质的积累,从而间接影响植物C 累积[37]。N 和P 共同参与了植物体内光合代谢和矿质代谢过程,N+P添加处理同时为植物提供了大量的必需元素,有利于植物在生长代谢过程中吸收更多的养分,导致C 储量增加。
Elser 等[38]曾报道,N和P同时添加在各种类型生态系统中都有较强的正协同效应,本研究也发现,在低、中种植密度情况下,N+P添加处理对幼苗单株及叶片C 储量的促进效果大于单一N或P添加处理。原因可能是仅添加N 会提高土壤的N︰P质量比,加剧了生态系统的P匮乏,从而限制树木生物量的增加[39]。一方面,N 添加使土壤酸性增强,促进土壤中的固相磷酸盐分解,并使其被土壤中的金属阳离子固定而沉淀,降低了P的有效性[40]。N 添加还会引起土壤中铝离子的溶出增加,抑制了植物对P的吸收[41];另一方面,本试验地为南方红壤区,土壤缺P较严重[42],因此土壤中供应的P不足以平衡N,单一N 添加处理会进一步增加P的匮乏程度,从而使P成为树木生长发育的限制因子[43]。P添加有利于提高生态系统中N 的利用效率,N+P添加处理对净初级生产力的促进效果更为显著[44]。本研究中的N+P添加处理,为樟树幼苗提供了生长所需的N 素,同时缓解了P限制,因此对幼苗碳储量的促进作用更强。
植物C的分配转移模式是植物种群提高竞争能力、克服逆境的重要策略[45]。研究表明,高密度的种植条件会抑制林冠的生长发育,在竞争压力较大的环境下,植物会优先将C分配给茎。本研究中随着种植密度增大,幼苗对叶的C分配比例减弱,而增加了对茎的分配比例。可能是种植密度的增大加剧了植物之间的竞争,茎对促进植物生长发育的作用增加,而叶片的作用相对减弱。另外,植物通过激素抑制侧芽的生长,将C从叶向茎转移,从而引起植物不同器官C储量的分配比例发生改变[46]。同时,由于高密度种植条件下植物的生长空间拥挤狭小,且林冠郁闭早,限制了树冠的生长发育,这也可能导致幼苗叶的C储量分配比例降低[47]。
本研究表明,种植密度和N、P添加处理的交互作用对幼苗枝和叶的单位面积C 储量,以及幼苗单株及枝和叶的C储量有显著影响。按照最优化分配理论,当光资源受限时,植物增大枝叶的分配;而土壤养分受限时,植物增大根的分配[48]。N、P添加处理增加了土壤养分,能够在一定程度上缓解高密度条件下的养分限制,在养分已经满足了樟树幼苗需求的情况下,光合作用成为幼苗生物量积累的限制因素,因而幼苗将更多的生物量向叶片分配,而叶片生物量需要更大的枝条来支撑[49],从而有利于幼苗枝和叶C 储量及单株幼苗总C储量的增加。随着种植密度的增大,植物个体对N、P等土壤营养元素的竞争加剧,同时林冠层郁闭较早,从而阻碍了林冠的生长,幼苗生长空间受到限制,树木的侧枝数量和长度均下降,这也会导致枝、叶和单株幼苗总C储量的减少[46,50]。
