中国水牛乳蛋白基因型与酪蛋白胶束粒度的关联研究
2020-01-16赵烜影刘建新任大喜
陈 晨,赵烜影,郭 鸰,李 玲,刘建新,任大喜*
(1.浙江大学奶业科学研究所,浙江杭州 310058;2.东北农业大学食品学院,黑龙江哈尔滨 150030;3.广西水牛研究所,广西南宁 530001)
中国养殖水牛超过1 200 万头,水牛乳产量居世界第三。与荷斯坦牛乳相比,水牛乳营养丰富,能够为人类营养发挥重要作用,但长期以来,我国对乳品的基础研究不足,对水牛乳、羊乳和骆驼乳等新兴乳品研究更加匮乏。酪蛋白是乳品中的主要蛋白成分,也是食品加工中最主要的原料成分之一,研究酪蛋白亚型结构对提升水牛乳品质起到重要作用。
自1955 年,Aschaffenburg 等[1]等第1 次揭示β-乳球蛋白(β-LG)的多态性以来,相继发现了αs1-酪蛋白(αs1-CN)、β-酪蛋白(β-CN)、κ-酪蛋白(κ-CN)的多态性。乳蛋白基因多态性与牛乳成分、泌乳性能以及乳制品加工特性等密切相关,因此国内外研究者开展了大量研究。前期研究发现,中国荷斯坦奶牛的κ-CN、β-CN 和β-LG 均存在多态性,且这些乳蛋白基因多态性与泌乳性能存在显著关联[2]。此外,还发现干酪凝乳性能和终产品品质与κ-CN 的B 基因存在正相关,与E基因存在负相关[3]。
在牛奶中,酪蛋白多以分散的、大致呈球形的胶体颗粒形式存在,被称为“酪蛋白胶束”,大小在150~230 nm 变化[4-6]。部分学者认为酪蛋白胶束大小的变化似乎与奶牛的年龄、产奶量、泌乳阶段或奶中的脂肪或蛋白质含量不相关[6],但酪蛋白胶束与干酪加工过程中的凝乳强度、干酪的硬度等性质密切相关[7]。关于胶束的精细结构及其如何受到酪蛋白遗传变异体的影响,目前仍没有明确共识[8-9]。酪蛋白组成与粒径之间是否存在相关性尚存争议,原因之一在于不同作者在酪蛋白胶束的制备和成分分析方面有差异,以及不同作者对胶束尺寸“大”和“小”的定义并不一致[10]。大多数对于酪蛋白胶束功能性遗传学的研究都集中在κ-CN变体、蛋白质粒度和牛奶功能特性之间的关系上[6]。即使有研究发现β-CN 的A2A2基因型与不良凝固性质相关,关于β-CN 亚型与蛋白质粒度的关联研究仍是较少的[11-13]。本研究拟采用反相高效液相色谱法来分析水牛乳中主要蛋白的多态性,并建立酪蛋白各亚型和酪蛋白胶束粒度的关联,以期为我国水牛品种改良和水牛乳质量的遗传选择提供新的见解,并促进加工技术转型升级以提高加工效率和乳制品质量。
1 材料与方法
1.1 实验材料
1.1.1 样品采集 水牛乳样采自广西水牛研究所牧场。2017 年3 月选取160 头泌乳中期的中国水牛,每头采集50 mL 新鲜乳样,4 000 ×g 离心15 min 除去乳脂肪,各取500 μL 分装于离心管中用于分析多态性,-20℃冻存备用,其余样品马上进行粒度分析。
1.1.2 试剂 乙腈、三氟乙酸(TFA)、甲醇均为色谱纯试剂,Bis-Tris、盐酸胍、二硫苏糖醇(DTT)、柠檬酸钠均为分析纯试剂。实验所用水均为Millpore 超纯水。αs-CN、β-CN、κ-CN、α-LA 和β-LG 标准品均购自sigma 公司。
1.1.3 仪器 安捷伦1260 型液相色谱(Agilent Technologies Inc 1260-LC)、离心机(上海沪湘仪 TW16-WS)、超声波清洗器(昆山市超声仪器有限公司 KQ-250DE)、涡旋混合器(上海精科实验有限公司 XW-80A)、超纯水仪(Millpore Direct-Q8 UV),Bettersize 2000 型激光粒度分析仪(丹东百特仪器有限公司)。
1.2 实验方法
1.2.1 多态性分析 向分装的冷冻乳样中加入等体积的工作液Ⅰ(0.1 mol/L BisTris,6 mol/L 盐酸胍,5.37 mmol/L柠檬酸钠,19.5 mmol/L DTT 的混合溶液),样品融化后振荡10 s 混匀,室温静置1 h。将混合溶液16 000 ×g 4℃离心5 min,除去表层乳脂,取底层溶液300 μL 于新的离心管中,加入工作液Ⅱ(以流动相A 为溶剂的4.5 mol/L盐酸胍溶液)稀释(体积比1:3),混匀,过滤到进样瓶中待测。
采用安捷伦1260 型液相色谱仪,色谱柱为C8 柱(300SB-C8),流动相A 为0.1% TFA 水溶液,流动相B 为0.1% TFA 乙腈溶液,检测波长214 nm,柱温45℃,进样量10 μL,流速0.5 mL/min。乳蛋白分析的梯度洗脱程序参照刘亚楠等实验方法[14]。
1.2.2 粒度测定 取40 mL 乳样1 500 ×g 15 min 4℃反复离心2 次,除去乳脂。通过Bettersize 2000 型激光粒度分析仪测定其粒度数据体积动量平均值(D[4,3]),面积动量平均值(D[3,2])和中位径(D[50])。
1.2.3 统计分析 根据标准品出峰时间,判定样品各个乳蛋白的出峰顺序。依据Bonfatti 等[15]液相出峰时间判定实验中乳蛋白多态性。收集各乳样的粒度数据采用SAS(9.2)软件的MIXED 程序对实验结果进行统计、分析。利用最小二乘法拟合线性模型,比较不同复合基因型牛乳的酪蛋白胶束粒度的差异。由于水牛场系谱数据不够全面,模型中不考虑水牛个体之间的亲缘关系,所有环境因素也因此剔除,使得基因效应不被环境变异所掩盖。所用模型:
式中,Y 为酪蛋白粒度;μ 为群体均值系数;Gi为复合基因型固定效应;H 为牛群固定效应;Pm为胎次固定效应;eijk为随机残差效应。
2 结果与分析
2.1 乳蛋白多态性分析 根据标准品的出峰时间,可以获得不同乳蛋白的出峰顺序。如图1-A 所示,6 种乳蛋白的出峰顺序为:κ-CN、αs2-CN、αs1-CN、β-CN、α-LA和β-LG[2]。αs1-CN 存在A、B 2 个等位基因,有AA、AB 和BB 3 种基因型,β-CN 存在A 型和B 型2 种多态性,κ-CN 有A 型、B 型、C 型和D 型4 种多态性。α-LA 存在A 型和B 型2 种多态性,αs2-CN 和β-LG 没有发现多态性。
2.2 乳蛋白多基因频率分析 水牛乳蛋白等位基因频率与各亚型统计结果见表1。其中αs1-CN 的2 种等位基因中B 的比例明显大于A,为81.56%,3 种基因型的频率中BB 最大,占66.87%。在β-CN 中,B 等位基因频率远高于A,3 种基因型的频率BB 最高,为64.38%。κ-CN 的4 种等位基因中,B 也是最高,为84.40%。α-LA 同样存在A 和B 2 种等位基因,B 频率远远高于A,占90.62%,BB 也高于AA。αs2-CN 和β-LG只存在BB 基因型。
本研究中,αs2-CN 和β-LG 不存在基因多态性,共得到7 种αs1-β-κ-CN 酪蛋白复合基因型。其中,AA-BB-BB 复合基因型个数最多,为46 个。AB-BBAB 和AA-AA-BB 复合基因型个数最少,仅为4 个。统计结果见表2。
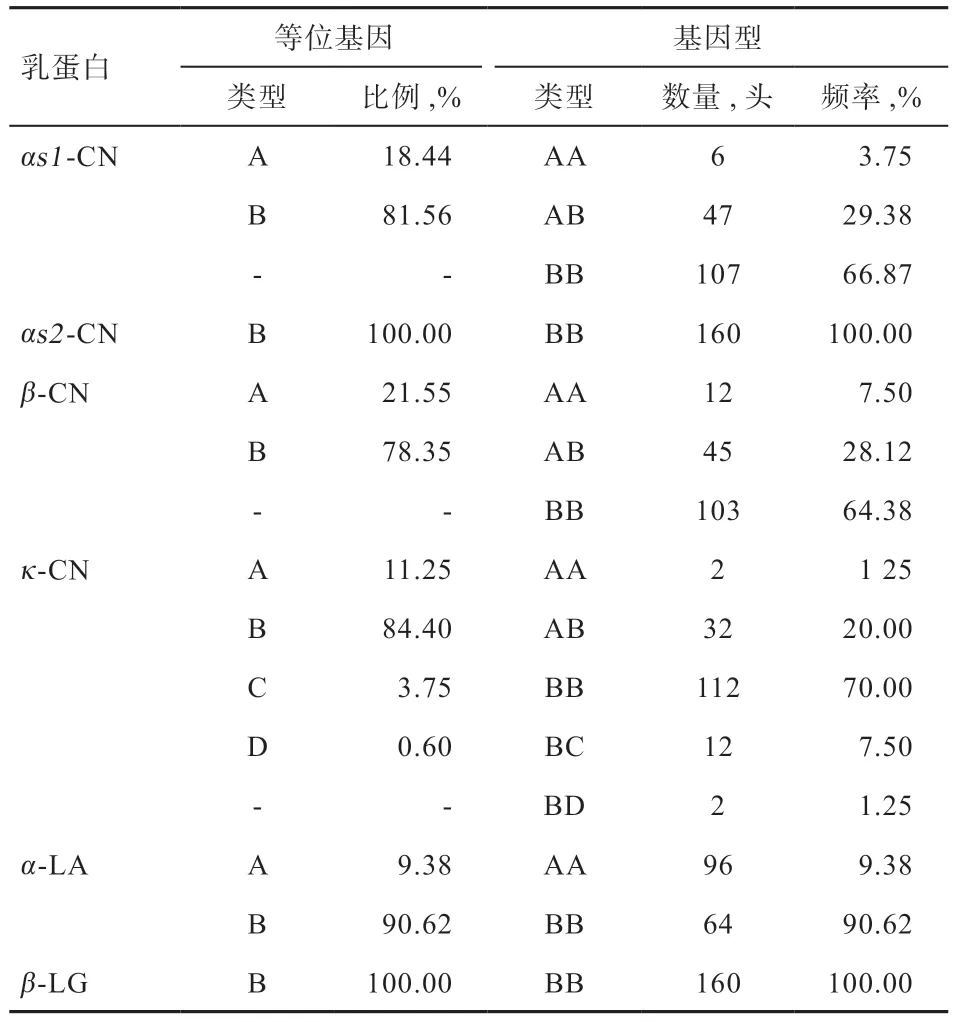
表1 水牛乳蛋白等位基因频率与基因型统计结果

表2 αs1-β-κ-CN 复合基因型
2.3 水牛乳酪蛋白粒度分析 本研究通过激光粒度仪分析水牛乳样品的蛋白粒度(D),结果如图2 所示,蛋白的表面积平均径D[3,2]分布于0.200~0.279 μm,主要集中在0.220~0.239 μm,约占蛋白总量的56%。蛋白的体积平均径D[4,3]分布于0.220~0.900 μm,其中0.240~0.249 μm 与0.250~0.259 μm 2 种不同 体积平均径的组分比例最高,分别占蛋白总量的20% 和22%。蛋白的中位粒径D50 主要集中在0.240~0.259 μm,约占蛋白总量的59%。
2.4 水牛乳酪蛋白与蛋白粒度的关联 利用SAS9.2 软件统计分析各酪蛋白与蛋白粒度的关联,结果如表3所示,κ-CN 的BB、AB 和BC 3 种基因型在D[3,2]上差异显著,并且AB 与BC 组内差异显著;κ-CN 的BB、AB 和BC 3 种基因型在D50 上差异显著,AB 与BC、BB 与BC 组内差异显著。通过D50 可以看出:拥有κ-CN 的C 等位基因的样品蛋白的中位粒径<拥有B等位基因的样品<拥有A 等位基因的样品。
αs1-CN 的AA、AB 和BB 3 种基因型在D[3,2]上有显著差异,并且AB 与BB 组内差异显著。β-CN的3 种基因型在蛋白粒度的3 项指标中差异不显著。

表3 水牛乳蛋白基因多态性与酪蛋白胶束粒度(μm)的关系
水牛的αs2-CN、β-LG 和α-LA 基本上不存在多态性或者样本数很少。水牛多态性主要在αs1-CN、β-CN、κ-CN 这3 种蛋白上,而这3 种蛋白是牛乳中的主要酪蛋白,对乳品加工影响显著,因此选取αs1-β-κ-CN 复合基因型进行研究。通过SAS9.2 软件统计分析6 种复合基因型乳蛋白粒度与基因型的关联,结果如表4 所示,这6 种复合基因型在D[3,2]和D50 上有显著差异。αs1-β-κ-CN 复合基因型中的AA-BB-AB 的D[3,2]和D50 分别为0.241 μm 和0.257 μm,均为6 种复合基因型的最大值,而AB-BB-AB 的D[3,2]和D50 分别为0.216 μm 和0.234 μm,均为6 种复合基因型的最小值,可以推断在复合基因型中αs1-CN 为AA 型的蛋白粒度大于AB 型。

表4 水牛乳蛋白复合基因型多态性与酪蛋白胶束粒度(μm)的关系
3 讨 论
3.1 水牛乳酪蛋白多态性分析 目前测定乳蛋白多态性的方法包括分子水平和蛋白水平两大类,其中高效液相色谱法是一种快速准确的测定方法。本研究发现αs1-CN 存在A 和B 2 种基因型,与王强等[16]、Maria[17]的研究结果一致。本研究中αs2-CN 全部为BB 型,与其他报道相同[18]。任大喜等[19]通过焦磷酸测序和液相色谱结合分析发现我国水牛的β-CN 存在A、B 和C 3 种等位基因。本研究发现κ-CN 的多态性较为丰富,有A、B、C、D 4 种等位基因,这与Pipalia 等[20]、Dayem等[21]分别用印度水牛与埃及水牛仅发现B 型的研究结果不同,本研究出现两例BD 型,其中的D 基因型此前并未在水牛中被发现过,可进一步验证。本研究发现α-LA 有A 和B 2 种基因型,不同于Dayal 等[22]在Bhadwari、Mehsana、Surti 和Murrah 4 种水牛中发现的A、B、C、D 和E 5 种基因型;而β-LG 仅有B 型等位基因,这与Vohra 等[23]研究发现有A、B、C 3 个不同基因型的报道不一致。原因一方面可能在于奶牛品种不同,因为乳蛋白组成不仅受基因型影响,还与奶牛体况、环境、饲养管理方法等因素有关,另一方面是本研究样本数量有限,所检测到的牛乳蛋白基因型相对较少。
3.2 水牛乳酪蛋白亚型与蛋白粒度的关联 本研究通过激光粒度仪分析广西水牛乳样品,测得蛋白粒径的分布范围与李子超等[24]通过纳米粒度分析仪和扫描电子显微镜研究发现水牛乳酪蛋白直径大小分布于43.8~825.0 nm的结果相近,但本研究中蛋白粒度与他们研究中酪蛋白粒度以458.7~531.2 nm 为主有差异。这可能与两地引进的水牛品种不同以及所处不同的泌乳阶段等因素有关。
本研究通过分析不同酪蛋白亚型与蛋白粒度的关联,发现κ-CN 的AB、BB 及BC 3 种亚型在D[3,2]和D50 上有显著差异,等位基因为A 的样品蛋白粒度<含B 的样品<含C 的样品。这与Day 等[10]研究荷斯坦牛乳发现的大酪蛋白胶束样品中只含有κ-CN 的AA 亚型,而在小酪蛋白胶束的乳样中发现了κ-CN 的AA、AB 和BB 亚型结果相似。而Bijl 等[5]研究发现,与AB 和BB 亚型相比,κ-CN 的BE 亚型与大的酪蛋白胶束相关,与本研究结果略有差异,可能是因为牛的品种不同。Pirisi 等[25]发现绵羊乳中含有αs1-CN 的蛋白胶束,平均粒径为DD 型< DC 型< CC 型,且三者之间有极显著差异,这与本研究发现αs1-CN 的AA、AB和BB 3 种亚型在D[3,2]上有显著差异的结果一致。本研究中水牛乳αs1-CN 的样品蛋白粒径AA 型< AB型<BB 型。Ketto 等[26]研究发现挪威红牛拥有αs1-CN的BC 亚型的蛋白胶束粒径小于BB 亚型,β-CN 的基因多态性对蛋白胶束粒径没有影响,这与本研究发现β-CN 各亚型在蛋白粒度上没有明显差异的结果一致。Day 等[5]研究荷斯坦牛乳发现的大蛋白胶束乳样只含有A1A1、A1A2和A2A23 种β-CN 亚型,而在小胶束乳样品中鉴定出包括B 和I 的遗传变体在内的相当多的表型混合物。Lodes 等[27]发现与含有A1A1基因型的牛奶相比,含有BB 基因型的β-CN 的牛奶具有更小的胶束尺寸,但在A2A2的情况下则不然,这与本研究结果不一致,可能是因为水牛的β-CN 亚型远少于荷斯坦牛的β-CN 亚型。
3.3 水牛乳复合基因型与蛋白粒度的关联 本研究发现这6 种复合基因型在D[3,2]和D50 上有显著差异。αs1-β-κ-CN 复 合 基 因 型 中 的AA-BB-AB 的D[3,2]和D50 均为6 种复合基因型的最大值,AB-BB-AB 的D[3,2]和D50 均为6 种复合基因型的最小值,并且AA-BB-AB 同AA-AB-BC 以及AB-BB-AB 组间差异显著。这与Ketto 等[26]研究挪威红牛发现αs1-β-κ-CN 复合基因型对酪蛋白胶束大小有极显著影响的结果一致,他们在BB-A1A2-BE 复合基因型中发现较大的蛋白胶束粒径。Gustavsson 等[28]研究发现β-κ-CN 的A1A2-AB复合基因型中的酪蛋白胶束粒径小于A2A2-AA 和A1A1-EE 复合基因型。这些研究都表明酪蛋白复合基因型对酪蛋白胶束大小有显著影响。
目前,国内外尚未有关于水牛乳蛋白多态性与胶束粒度关联的报道,前人研究表明酪蛋白胶束与干酪加工等密切相关。水牛乳是加工马苏里拉等干酪的优质原料,因此本研究结果可为特定水牛品种的选育提供指导,为今后水牛乳的深加工提供支持。
4 结 论
本研究采用反向高效液相色谱法分析了我国南方地区水牛乳蛋白多态性,结果发现水牛的κ-CN、β-CN、αs1-CN 和α-LA 都 存在 多 态性,而αs2-CN 和β-LG 只有1 种基因型,其中κ-CN 发现了一种新的等位基因D,揭示了单个牛奶样品中酪蛋白基因型与蛋白胶束大小的关联,发现κ-CN 以及αs1-CN 各亚型在乳蛋白D[3,2]上有显著差异,等位基因为A 的蛋白粒度要大于其他等位基因,筛选出的6 种αs1-β-κ-CN 复合基因型在乳蛋白D[3,2]以及D50 上有显著差异,AB-BB-AB 的D[3,2]和D50 均为6 种复合基因型的最小值。研究结果表明乳蛋白多态性与酪蛋白胶束粒径存在显著关联,为今后水牛品种的改良和原料乳品质提高提供了基础数据。
