犬体型大小相关基因的研究进展
2020-01-16韦云芳李飞翔毛爱国陈方良万九生
韦云芳 ,李飞翔,汪 斌,李 静,毛爱国,陈方良,万九生,陈 超*
(1.公安部昆明警犬基地,云南昆明 650204;2.警犬技术公安部重点实验室,云南昆明 650204;3.云南农业大学动物科学技术学院,云南昆明 650201)
家犬是人类驯化历史中独一无二的杰作。许多世纪以来,没有其他哺乳动物能像犬一样,与人类产生如此紧密的联系,更没有如此显著的表型差异。当今所有家犬都是距今约15000 年前由灰狼驯化而来[1]。犬被驯化之后,人类根据自己的意愿和使用目的加大了对犬的选育强度,由此形成了如今形态各异的现代犬种。目前,世界上有超过400 多个不同品种的犬[2-4],除了金鱼,犬是与其野生祖先表型差距最大的驯化动物。然而,现代家犬品种形成的时间并不长,大概是200~300 年前,这也说明了人工选择的显著效果[3]。现今每一个家犬品种都有着严苛的品种标准以维持其固定的表型,这些标准将持续地对某一品种犬施加选择压力,这种压力降低了品种内的表型多样性和遗传异质性,但增加了品种间的巨大差异性。体型是犬最显著的表型性状,家犬是所有陆地脊椎动物中体型差异最大的动物,犬的体型差异之大,堪称各物种之最[5]。最小品种犬与最大品种犬的骨骼大小相差大概40 倍。吉娃娃的平均体重只有1.8 kg,而獒犬体重高达90 kg,几乎是最小的吉娃娃的50 倍[4-6]。据“吉尼斯世界纪录”记载,世界上最高的犬是一条名叫“宙斯”的大丹犬,其身高为111.8 cm,体重达70.3 kg,直立时身高高达2.25 m。最小的狗是一只名叫“米莉”的吉娃娃,体重453 g,身高只有9.65 cm(http://www.guinnessworldrecords.com/)。那么,同样是灰狼的后代,在相对短暂的时间内,犬是如何演化出这么多体型迥异的品种?本文就犬体型性状相关基因IGF1、IGF1R、GHR1、GHR2、SMAD2、STC2、HMGA2、IGSF1、ACSL4及IRS4的研究进展进行综述。
1 犬体型大小的遗传研究
针对犬体型大小的第一个研究是从葡萄牙水犬(Portuguese Water Dog,PWD)开始的[7],这是因为在美国葡萄牙水犬是一个相对较新的品种,可以追踪24 代,一直到当初的31 个建群者。而且葡萄牙水犬的品种标准对其体型大小的界定允许有2 倍的差异,因此导致该品种内的个体体型差异较大,加上具备详细的系谱记录,使它成为体型性状研究中最好的试验对象。Chase 等[7-8]对500 只葡萄牙水犬的X 射线骨骼图进行分析,制定了92 个骨骼参数,利用全基因组微卫星扫描数据,结合葡萄牙水犬种群骨骼变异的主成分分析(Principal Component Analysis,PCA),发现15 号染色体(CFA15)上一个15 Mb 的区域有2 个数量性状位点(QTL)(FH2017 和FH2295)与葡萄牙水犬的体型大小密切相关。
2007 年,Sutter 等[5]对决定家犬体型大小的基因位点的确定成为寻找性状决定位点的经典工作之一。该研究仍是从葡萄牙水犬开始,并把研究对象扩大到143个品种、3241 只犬,首先对15 号染色体15 Mb 区域的SNP 进行重测序,进一步将该区域锁定到4 Mb 以内,然后研究者对463 只葡萄牙水犬在这个区域的116 个SNPs 和体型大小进行关联分析,发现在胰岛素样生长因子1(Insulin-like Growth Factor 1,IGF1)基因附近出现了峰值,而IGF1也是影响人和小鼠个体大小的重要候选基因。然而由于葡萄牙水犬品种内个体在IGF1区域内高度连锁,确定具体的位点非常困难,因此研究者对来自于526 只犬的116 个SNPs 进行关联性分析,其中有23 种小型犬(<9 kg),20 种大型犬(>30 kg)。在此,研究者发现在IGF1区域,杂合率降低,大型犬和小型犬出现了明显的遗传分化,而小型犬的平均杂合率只有大型犬的25%,因此进一步确认了此区域是大、小体型的犬分化的主要关联区域,进一步数据分析将区域锁定到了20 个SNP,最终确定了5 号SNP(CFA15,44,228,468)是最佳候选对象,小型犬中的碱基是A,而大型犬中的碱基是G。进一步对143 种品种犬中等位基因A 的频率进行分析,发现A 的频率和品种的体型强烈负相关,以此确定了5 号SNP 是家犬体型大小的因果突变。因此,IGF1基因上单个SNP 的突变决定了犬的小体型。
随后一个很有开创性的工作是Jones 等[9]率先使用各犬种的品种标准来定义表型与基因标记关联分析。因为现代犬种是由严格的品种标准所定义的,品种标准描述了某个品种中个体的理想体型特征。对于体型这样的强选择性状的遗传研究,品种标准可以用作个体遗传决定表型的代表,由此开启了在犬类形态学的遗传研究中使用品种标准进行分析的方法,而不是对每只犬进行个体测量,极大地简化了相关工作。
后来,随着“CanMap project”的开展,又相继确认了多个与体型大小相关的重要基因座。2012 年,Hoopes 等[4]专门针对平均体高在25 cm 以下的915 只小型犬进行研究,在3 号染色体(CFA3)上鉴定了一个新的犬体型相关基因座——胰岛素样生长因子1 受体(IGF1R)基因。2013 年,Rimbault 等[10]通过精细定位发现了对体型有显著影响的另5 个基因,分别是生长激 素 受 体(Growth Hormone Receptor,GHR)GHR1和GHR2基因、高迁移率族蛋白A2(High Mobility Group AThook 2,HMGA2)、斯钙素2(Stanniocalcin 2,STC2)、SMAD 家族成员2(SMAD family member 2,SMAD2);此外,46%~52.5%犬种的体型差异可以通过包括IGF1和IGF1R在内的6 个候选基因上的7 个基因座来解释。在标准体重小于41 kg 的品种中,这6 个基因的突变可解释64.3%的犬体型变小的原因。2017 年,Plassais 等[11]使用Illumina 公司Canine HD 芯片(170000 SNPs)对大型犬种与小型犬种进行了一项全基因组关联分析(Genome-Wide Association Studies,GWAS),发现大体型犬的骨骼大小和体重与胰岛素受体底物4(Insulin Receptor Substrate,IRS4)、长链酯酰辅酶A 合成酶家族成员4(Acyl-CoA Synthetase Long-chain family member 4,ACSL4)和免疫球蛋白超家族成员1(Immunoglobulin superfamily member 1,IGSF1)基因的变异密切相关。
对犬体型大小的相关重要研究总结见表1。伴随着技术的不断发展,体型大小相关基因的发现及研究结果不断完善。随着全基因组高密度SNP 芯片技术的发展,SNP 分型的规模越来越大,从60968 发展到了170000。另外,研究手段和分析方法不断丰富,包括精确定位(Fine-Mapping)、全基因组测序(Whole Genome Sequencing,WGS)及全基因组关联分析(GWAS)等方法的综合运用。再者就是研究材料不断丰富,从1 个犬种(葡萄牙水犬,PWD)330 头犬到88 个犬种855 头犬,甚至是148 个犬种2801 头犬。
2 犬体型大小相关基因的研究
目前,研究发现10 个候选基因的变异决定了大多数犬体型的大小(表2)。前3 个基因的遗传变异决定了大体型犬的体高和体重。后7 个基因大概可以解释≤41kg 的中小型犬的60%体型差异。
2.1IGF1和IGF1R基因 IGF1 是介导生长激素作用的一种单链多肽,也被称作促生长因子(即Somatomedin C),是胰岛素样生长因子家族(IGFs)的重要成员,主要产生于肝脏细胞,通过内分泌、旁分泌或自分泌途径作用于靶器官,主要与IGF1R 结合从而刺激下游多条信号转导途径,将信号转导至细胞核内,激活并释放转导因子,进而对哺乳动物的细胞增殖和分化及凋亡、机体的生长发育、组织修复、营养代谢等方面起着重要的调控作用[14]。
IGF1 和IGF1R 都是动物生长轴IGF1 通路上的重要因子,具有显著的促生长和分化作用,在胚胎发育和个体生长中起着重要作用,并且该通路与生长激素(Growth Hormone,GH)通路之间存在复杂的相互关系。许多研究都发现IGF1 通路在控制犬体型大小方面起着重要作用。Sutter 等[5]鉴别出IGF1是决定犬体型变小的关键基因,且具有IGF1基因变异的小体型犬的血清中IGF1激素水平也显著低于大体型犬。后来,IGF1R基因也被证明是影响犬体型大小的重要基因[4]。当IGF1和IGF1R基因突变时,会抑制胰岛素信号通路功能的发挥,从而影响个体大小。在小鼠和人上,都已证实IGF-1和IGF1R基因变异会导致生长发育迟缓及体型、重量的减小[15-17]。目前,畜禽IGF1 的研究主要集中在IGF1基因的克隆、结构与功能、多态性及其与动物生产性能和经济性状的关系等方面。
2.2GHR1和GHR2基因 生长激素(GH)是由脑垂体前叶分泌的一种蛋白激素,在动物生长发育及代谢中有重要作用。GH 首先是与靶细胞膜表面的GHR 结合,启动细胞内信号传导,促进IGF1 的表达,再通过血液循环到达生物体局部组织,促进细胞的新陈代谢过程[18]。GHR属于细胞分裂素/血细胞生成素受体超家族(Cytokine/Hematopoietin Receptor Superfamily),是一种特异、高效的膜蛋白,其在细胞内的表达量对GH 发挥作用有重要影响,从而影响生物体的生长发育,如性连锁矮小鸡的生长迟缓症状就是因为GHR基因突变导致动物组织中GHR 数量显著减少甚至缺乏所致[19-20]。GHR基因是第一个被证明与生长有关的基因。人类中,GHR基因突变导致的GHR 结构与功能异常都会抑制GH 发挥正常生理作用,最终造成生长发育不良、迟缓和身材矮小、侏儒症等[21]。在犬上,GHR1和GHR2基因的变异都会对犬体型大小造成影响[10]。有研究表明,猪体内GHR 的失活也会导致成年猪体重和体尺的减少[22]。

表1 犬体型大小相关研究
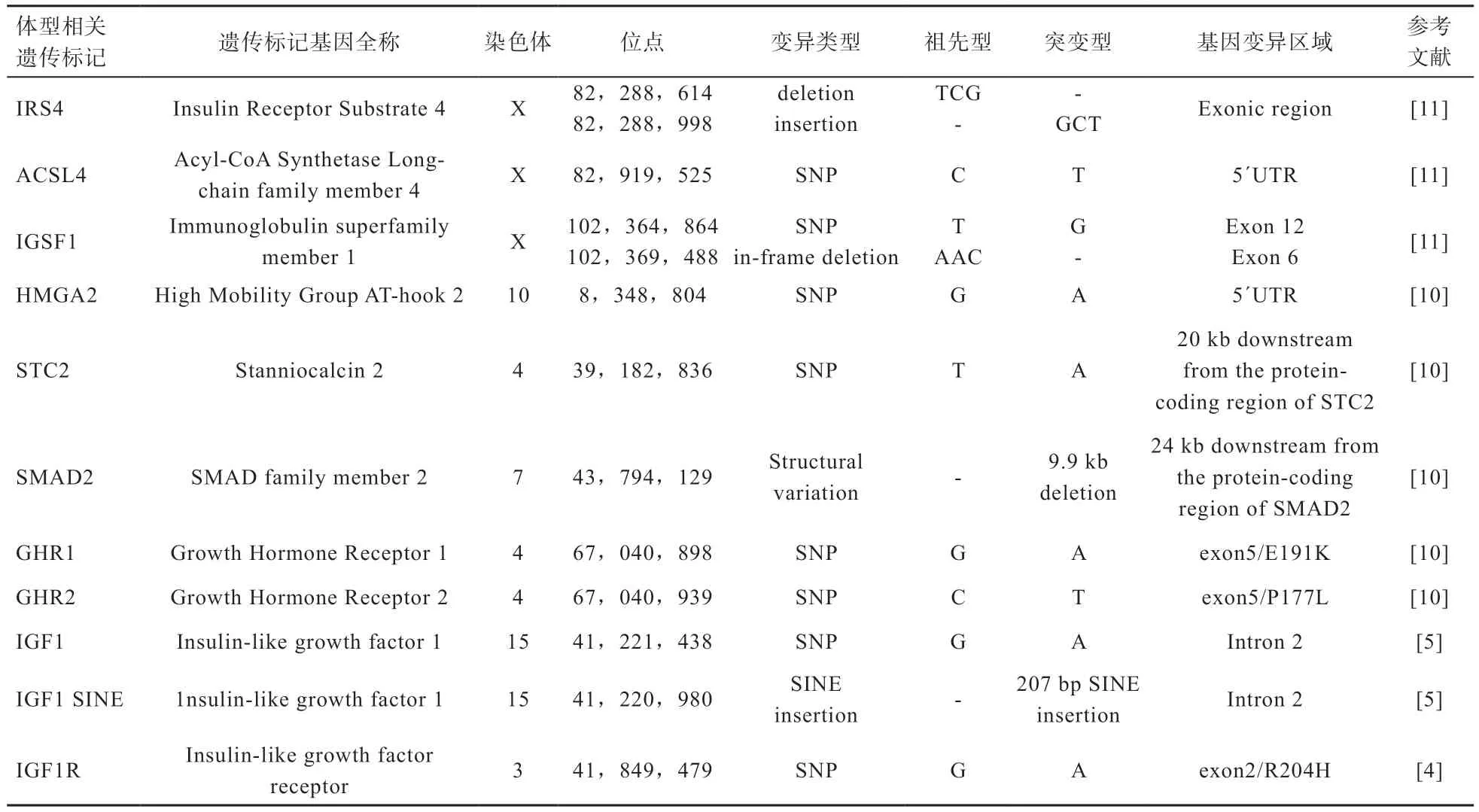
表2 犬体型大小相关候选基因的信息
2.3HMGA2基因 HMGA2 属于高迁移率蛋白A 家族中的一员,是一种非组蛋白染色体蛋白,本身缺乏转录活性,但能通过与染色质结合而改变其结构,或者直接与相关蛋白结合发生作用,继而调节其他基因的转录,从而影响胚胎形成、组织发育、生长调节及肿瘤发生等过程[23]。小鼠上,HMGA2的1 个等位基因的失活会造成小鼠体重减少20%,2 个等位基因都失活则会使小鼠体重减少60%[24-25]。相反,HMGA2过表达导致小鼠出现巨人症和脂肪瘤[26]。HMGA2 蛋白在小鼠胚胎发育和细胞分化增殖中也发挥着重要作用,在胚胎期以及不成熟组织中大量表达,而在分化成熟的组织中几乎不表达[27]。在人类中,HMGA2基因的变异会导致个体身材矮小[28]。在猪上,HMGA2基因敲除猪出现侏儒症表型,特征是出生体重过轻、生长迟缓及体重减少[29]。此外,HMGA2基因缺失影响了猪胎儿在母体子宫内接受的资源,若是子宫内同时存在正常与HMGA2-/+、HMGA2-/-等个体,则HMGA2-/-个体无法在怀孕期间存活,且HMGA2-/-与子宫绒毛接触不良,显示子宫跟胎盘间的连结较差;若子宫内只存有HMGA2-/-,这些胎儿就能存活并正常发育[29]。另外,目前已知HMGA2基因表达水平会影响多种动物的体型,包括犬[9-10,30]、兔[31]、马[32]等多种动物。最近的研究还表明,HMGA2基因变异能影响鸟喙的大小[33]。
2.4STC2基因 STC2 属于STC 家族中的一员,是哺乳动物器官及组织中广泛表达的一种分泌性的糖蛋白激素,参与调节多种生理及病理过程[34-35]。STC2 通过与妊娠相关血浆蛋白-A(PAPP-A)共价结合从而抑制PAPP-A 的蛋白水解活性,进而阻止胰岛素样生长因子蛋白4(IGFBP4)的裂解,由此抑制胰岛素样生长因子(IGFs)的活性而阻碍哺乳动物的正常生长[36]。在小鼠上的研究表明STC2 能有效调节小鼠出生后的生长。STC2基因过表达的小鼠在体型减小了45%[37],而STC2基因敲除小鼠比同窝出生的野生型小鼠要大15%[38]。在犬上,Rimbault 等[10]研究发现STC2基因的变异会导致犬体型的变小。而关于人类身高的研究表明,STC2基因的2 种不同变异会对人类身高产生影响[39]。
2.5SMAD2基因 SMADs 是一类新的细胞内信号转导蛋白家族,能够转导来自转化生长因子β(Transforming Growth Factor-β,TGF-β)的信号因子,其家族成员与转化生长因子TGF-β共同调节细胞的增殖、分化、凋亡及间质的合成,从而在生物体各器官组织的生长发育过程中起着极其重要的作用。在成人中,SMADs家族成员还负责组织修复和免疫调节。SMAD2基因是SMADs 蛋白家族的成员之一,属于受体激活型SMADs,是转导TGF-β信号的重要胞浆内信号级联分子,根据转化生长因子TGF-β超家族成员的信号,在细胞膜与细胞核之间来回移动,将TGF-β信号直接由细胞膜转导入细胞核内,从而调节特定靶蛋白的转录[40]。SMAD2 功能失活或表达过低可能影响TGF-β的信号转导并参与肿瘤的形成。
2.6IRS4基因 Plassais 等[11]首次发现IRS4基因与犬的大体型密切相关,但在人上并没有发现身高与IRS4基因有任何相关性。IRS4属于胰岛素受体底物(The Insulin Receptor Substrate,IRS)分子家族最新成员,该基因编码胰岛素受体底物-4,于1997 年由Lavan 等[41]成功克隆,是胰岛素胞浆内信号传导的关键分子之一[42],在下丘脑组织中高度表达[43]。IRS4 分子是胰岛素受体的直接作用底物,被磷酸化后,促发胰岛素生理信号在胞浆内的逐级传递过程,激活葡萄糖转运蛋白4(GLUT4)等下游效应分子[44]。另外,它参与GH/ IGF-1 途径并与多种生长因子受体(如IGF1R)相互作用,增强IGF1的表达从而刺激细胞的生长[45]。IRS4基因也受雌激素调节[46],这可以部分地解释雌激素和身体脂肪分布之间建立的联系[47]。Fantin 等[43]建立了IRS4基因敲除小鼠模型(IRS4-/-),发现IRS4敲除小鼠在生长、繁殖和糖代谢方面均出现轻度缺陷,说明IRS4 是参与机体生长与发育及糖代谢调节的信号分子。在人类研究中,IRS4基因多与Ⅱ型糖尿病、代谢紊乱、精神分裂症、肥胖等疾病相联系[48]。还有研究表明IRS4 会耦合到瘦素受体上,而瘦素和胰岛素都能够通过作用于下丘脑神经元调节生物体内的能量消耗和葡萄糖稳态,从而参与控制食物摄入,长期控制肥胖,促性腺激素的分泌以及繁殖性能的调控[49]。在猪上,IRS4基因的多态性可能引起脂肪沉积表型变异[50]。
2.7IGSF1基因IGSF1基因编码质膜糖蛋白,参与甲状腺激素的生物合成,并参与甲状腺激素途径[51]。据报道,在人类中,IGSF1基因的突变与X 连锁的IGSF1 缺乏综合征有关,IGSF1基因上的突变使其编码蛋白无法移动到细胞表面发挥正常功能,从而导致甲状腺功能减退,因此也与人类肥胖有关[51]。在小鼠上的研究也表明,缺乏IGSF1 的雄性小鼠显示垂体和血清促甲状腺激素(TSH)浓度降低,垂体促甲状腺激素释放激素(TRH)受体表达降低,体重增加[51]。在犬上,Plassais 等[11]研究发现IGSF1 与犬种的体高密切相关,在大型犬种中鉴定出2 种突变(单密码子缺失和错义突变)都位于IGSF1 蛋白的高度保守的免疫球蛋白样结构域。有意思的是,IGSF1基因还与犬的头型长短有关[12,52]。而在人上,患者身体上的某些缺陷包括面部的异常(如睑裂狭小、小头、唇腭裂等)和生殖器畸形都可能与IGSF1基因的短片段重复有关[53]。
2.8ACSL4基因 ACSLs 是脂肪酸代谢中至关重要的酶,在哺乳动物中已经鉴定出5 个该家族成员(ACSL1、3、4、5、6)。ACSL4 定位于细胞内膜,特异性结合长链多不饱和脂肪酸,参与脂肪酸的合成和分解代谢过程。研究表明,增加肝脏中ACSL4 的表达可促进脂肪酸的摄取[54]。Plassais 等[11]发现,ACSL4基因与大体型犬中发达的肌肉性状高度相关,但与矮胖的小体型犬健壮结实的肌肉性状无关,这表明在大体型犬中发现的ACSL4基因变异会大大增加犬的体型。在猪上,ACSL4控制猪肌肉和背部脂肪厚度,ACSL4的突变与背膘厚相关[55-56]。
对犬体型相关的候选基因的QTL 的遗传变异研究表明,重要候选基因的组合中突变型等位基因越多,犬的体型就越小。然而,其中某些基因在犬体型减小上似乎比其他基因有更强的效果。有研究表明,IGF1R、GHR2和HMGA2被认为具有更强的体型减小效应,具有较弱体型减小效应的是IGF1、GHR1和STC2[57]。
3 结语与展望
犬体型性状是一个复杂性状,受到多个基因的影响。目前,研究者已发现10 个基因的遗传变异对犬的体型大小有显著影响,这些遗传标记的确定为犬体型大小的预测提供了可能,通过遗传标记预测犬体型大小将大大有利于犬的健康,例如根据体型制定相应合理的饮食和运动计划。此外,预测犬体型的大小将有助于选育具有合适体型及与体型相关性状的理想个体。
虽然上述遗传标记可以解释大多数犬体型大小的变异,但是具体的影响机制尚未明确,仍需要进一步探索。随着上述候选基因在各种动物中的遗传变异与生长发育表型关系的深入挖掘和研究,这些分子标记在动物生长发育中的作用将越来越清晰,为理解和阐明人类及其他家养动物在体型上的遗传变异提供科学线索。另外,随着检测手段的升级、犬种及样本量的扩大,可能还会检测到与犬体型大小相关的其他候选基因和相关分子标记,并会在动物体型大小的遗传机制上有更多有价值的发现。
