GJB2基因突变在长岛型掌跖角化病发病中的作用
2020-01-10刘慧茹叶碧珍魏转弟
刘慧茹,叶碧珍,魏转弟
作者单位:东莞市中医院皮肤科,广东 东莞523000
长岛型掌跖角化症(Nagashima-type palmoplantar Keratosis,NPPK)是一种常染色体隐性遗传病,男女发病率相似,临床表现的特征为掌跖角化症,该病于1977年首次在长岛被报告[1]。其临床特征包括手掌和脚掌上界限清楚的红斑并轻中度角化过度;长岛型掌跖角化症通常在婴幼儿期开始发病,表现为轻微的掌跖斑疹病,皮疹进展慢[2]。
GJB2属于β-族间隙连接蛋白家族,按蛋白分子量命名为Cx26。间隙连接蛋白与缝连接蛋白组成一个完整的缝隙连接通道,在细胞间信息传导和物质交换起重要作用。表皮的基底层含有增殖的角质形成细胞,分化产生的细胞可迁移到棘突和颗粒层[3]。角质形成细胞具有许多细胞间连接,包括间隙连接[4]。β-族间隙连接蛋白突变可能诱导皮肤病发生,其中Cx26的GJB2突变会导致皮肤病综合征,包括耳聋的掌跖角化病(PPK),Vohwinkel综合征(VS)和角膜炎鱼鳞病耳聋综合征(KID)[5-7]。Cx31(GJB3)和Cx30.3(GJB4)突变引起的皮肤病症状相似,主要表现为变异性红皮肤角化症(EKV)[8]。目前大量研究集中在间隙连接蛋白导致KID综合征的作用机制,但对于与NPPK相关的突变鲜有研究。
本研究于2017年收集了家系的2例NPPK病人,对病人的外周血提取DNA,进行SERPINB7基因外显子8常见突变位点c.796C>T(p.Arg266Ter)进行测序,未发现突变。因此,我们推测该家系病人可能是由于其他相关基因(GJB2、GJB3、GJB4、GJB6、GJA1)突变引起。本研究拟通过鉴定NPPK的突变位点,阐明NPPK的发病机制,为NPPK的治疗提供一定的理论依据。

表1 长岛型掌跖角化症(NPPK)中GJB2、GJB3、GJB4、GJB6、GJA1基因PCR扩增引物序列
1 资料与方法
1.1 一般资料 研究对象为收集的2例家系NPPK病人,分别于3岁和4岁开始发病,临床表现为双手掌、指背、足底、手腕、踝部、跟腱等对称性红斑、角化性斑丘疹、斑块伴脱屑。组织病理学表现为表皮角化过度、颗粒层和棘层增厚,无表皮松解性角化过度的特点,真皮浅层少量毛细血管扩张,少量炎症细胞浸润。该例病人的临床及组织病理符合长岛型掌跖角化症。
1.2 实验方法
1.2.1 DNA提取与sanger测序 收集NPPK病人外周血3~5 mL,根据DNA提取试剂盒说明书提取血液DNA,利用紫外分光光度计检测DNA的浓度和纯度,保存-70℃备用。从美国国家生物技术信息中心(https://www.ncbi.nlm.nih.gov/)中 获 得 GJB2、GJB3、GJB4、GJB6、GJA1这5个基因的序列,设计引物(表1)。以下条件进行PCR扩增:95℃预变性3 min,95℃变性30 s,58℃退火30 s,在72℃延伸2 min,30个循环,72℃延伸10 min终止反应。1%琼脂糖凝胶电泳检测PCR产物,将获得的5个基因的PCR产物送华大基因进行正反向引物测序。用Gene tool软件将测定的序列与标准序列进行比对,标 准 序 列 参 考 :GJB2:NM_004004,GJB6:NM_001110219, GJB3: NM_001005752, GJB1: NM_000166,GJB4:NM_153212。
1.2.2 体外转录及卵母细胞显微注射 将Cx26和Cx31克隆到pCS2+表达载体中,用于非洲爪蟾卵母细胞的功能研究[9]。通过定点诱变制备Cx26-S183F结构体[10],并将DNA结构体克隆到pBlueScri1ptⅡ载体中并测序,然后亚克隆到pCS2+表达载体中。将定点诱变的结构体使用SP6 mMessage mMachine体外转录试剂盒线性化并转录。从非洲爪蟾雌性中收集卵母细胞,并在改良的Barth(MB)培养基中培养[11],向卵母细胞注射10 ng反义非洲爪蟾Cx38寡核苷酸[12],然后单独或组合注射连接蛋白转录物,注入水的卵母细胞作为阴性对照。Cx31 RNA和其他cRNA都以相同的浓度注射。
1.2.3 记录半通道电流 卵母细胞注射各种cRNA 24h后使用Gene Clamp 500放大器检测其半通道电流。将卵母细胞培养在不添加Ca2+的MB培养基中,在室温条件下进行膜片钳实验。将电极拉至1~2 MΩ的电阻,电极内灌注电极内液。通过记录半通道电流:钳制电压为-40 mV,电压从-30 mV去极化至+40 mV,阶跃10 mV每次持续5 s,从而获得半通道电流-电压(I-V)曲线。并在初始钳制电压-40 mV情况下,测量两个细胞的电流变化来计算连接电导(Gj)。一个细胞受±20 mV的交变脉冲,在另一个细胞中记录由电压变化产生的电流,其大小与连接电流(Ij)相等,通过Ij除以电压差计算电导率,即Gj=Ij/(V1-V2)。
1.2.4 蛋白质印迹法 卵母细胞如1.2.2做相同的处理,24 h后提取蛋白。在12%的SDS凝胶上电泳分离并转移到硝酸纤维素膜上,用5%的脱脂奶粉37℃封闭 1 h,用 CX26(GBJ2)或 Cx31(GBJ3)(ABCAM,Cambridge)的多克隆抗体进行检测,然后用辣根过氧化物酶标记的二抗37℃孵育1 h。用GADPH(ABCAM,Cambridge)作为内参。使用Image J软件对条带灰度值进行量化分析。
1.2.5 免疫共沉淀 卵母细胞如1.2.2做相同的处理,24 h后利用膜蛋白提取试剂盒提供的说明书提取膜蛋白。用PBS清洗两遍珠子,用PBS将Protein A agarose配制成50%浓度,蛋白样品中加入Protein A琼脂糖珠去除非特异性蛋白。用Cx26抗体孵育,加入Protein A琼脂糖珠来捕捉抗原抗体复合物,收集琼脂糖珠-抗原抗体复合物,样品煮沸5 min变性。在SDS凝胶上进行电泳,转膜后使用Cx26或Cx31的抗体进行WB检测蛋白质。
1.3 统计学方法 采用GraphPad Prism 6.0和SPSS 11.0软件对实验数据统计分析,进行单因素方差分析One-Way ANOVA,并用Student’s t检验进行差异比较分析,P<0.05为差异有统计学意义。
2 结果
2.1 sanger测序筛选出GJB2突变引起NPPK GJB2基因核苷酸序列第584位碱基由C突变成T(c.548C>T)发生了突变,导致编码的蛋白第183位的丝氨酸被苯丙氨酸取代(p.Ser183Phe)。NPPK病人的父母均为GJB2p.Ser183Phe杂合子突变携带者,100个与该家族不相关的正常汉族人中没有检测到p.Ser183Phe突变位点。
2.2 Cx26突变体无功能且抑制野生型连接蛋白Cx31 已有研究证实Cx31突变体可引起严重的皮肤病,当其与野生型Cx31共表达时,通道电导率降低,影响间隙连接通道的能力[13]。因此我们猜测Cx26突变体可能通过与Cx31互相作用从而出现NPPK。为证实这一点,Cx26-S183F及其他表皮连接蛋白在非洲爪蟾卵母细胞中表达及Cx31在非洲爪蟾卵母细胞中表达,检测间隙连接电导Gj。用水注射的卵母细胞阴性对照组的电导几乎可忽略(Gj=0.18 μS),而只注射Cx26的细胞的平均Gj为8.6μS。在单独注射Cx26-S183F的卵母细胞中测量的电导与阴性对照组细胞相同(Gj=0.13μS),说明功能丧失。当共同注射Cx26-S183F和Cx26时,平均电导为2.9 μS,显着低于单独注射Cx26的(P<0.05,图1A)。Cx26-S183F还显示出对Cx31的反式显性抑制,当两者都存在于卵母细胞中时,Cx26-S183F显著抑制Cx31(Gj=0.77 μS,P<0.05,图1B)。结果表明Cx26-S183F能有效抑制野生型Cx31,Cx26突变体与Cx31互作可能会产生NPPK。

图1 检测各处理组(注入水、Cx26、Cx26-S183F、Cx26-S183F+Cx26)卵母细胞的间隙连接的电导率:A为检测野生型Cx26、Cx26突变体及Cx26-S183F+Cx26的电导率,B为检测野生型Cx31、Cx31+Cx26及Cx31+Cx26-S183F的电导率
2.3 突变蛋白表达缺失不会引起电导率的降低用蛋白质印迹法检测野生型和突变体连接蛋白的表达。结果显示,注射Cx31、Cx31+Cx26、Cx31+Cx26-S183F的处理组中均检测到Cx31蛋白(31kDa)(图2A),灰度值分析表明各处理之间差异无统计学意义(P>0.05,图2B)。同理,在Cx26、Cx31+Cx26、Cx26-S183F、Cx26+Cx26-S183F及Cx31+Cx26-S183F处理组中均检测到Cx26(26kDa)(图2C),且表达量一致,定量分析显示各处理组之间差异无统计学意义(P>0.05,图2D)。说明Cx26-S183F突变体存在时功能活性的丧失与Cx31的翻译效率无关。
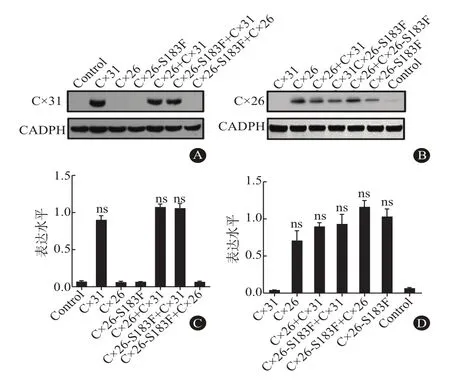
图2 蛋白质印迹法检测长岛型掌跖角化症(NPPK)突变体及野生型Cx31的表达水平:A为WB检测各处理组中Cx31的表达情况;B为WB检测各处理组中Cx26的表达情况;C为量化分析Cx31的蛋白表达量;D为量化分析Cx26的蛋白表达量
2.4 Cx26突变体与Cx31的互相作用 为了证实Cx26突变体和Cx31可能形成异型通道,我们对共表达Cx31和Cx26突变体的细胞进行免疫共沉淀实验。结果表明,在Cx26相关样品中检测到Cx26蛋白,而在含有Cx31的样品中均检测到Cx31蛋白(图3A和图3B),说明细胞裂解液可进行后续CO-IP实验。CO-IP显示Cx26蛋白在野生型和突变体样品中有表达,Cx26+Cx31样品在31KDa出现较弱的条带,说明野生型Cx26与Cx31结合较弱。共表达Cx31和Cx26-S183F样品中蛋白的表达量较高,表明Cx26突变体蛋白与Cx31发生免疫共沉淀(图3C)。
2.5 Cx26突变体不能形成功能性半通道 KID综合征突变体单独存在时通常形成活跃的半通道[14-16],而NPPK突变是否会出现类似的情况,为证实这一结果,在单个卵母细胞中表达Cx26和Cx26-S183F来分析半通道活性。结果显示,注射水的卵母细胞对照在所有电压阶段被限流(图4A),Cx26半通道活性在去极化时出现外向电流(图4B),与注射Cx26的细胞相比,Cx26-S183F的膜电流显著地降低(图4C)。以平均膜电流和膜电压作图,表达CX26的细胞出现较高的外向电流,并随着去极化的增加而增加,在+60mV时,Cx26产生的电流显著高于对照及Cx26-S183F突变体(P<0.05,图4D)。结果说明,注射突变体的细胞天然半通道活性丧失,Cx26-S183F与野生型Cx26的共表达导致半通道活性降低。

图3 免疫共沉淀(CO-IP)检测长岛型掌跖角化症(NPPK)中Cx26突变体与Cx31的互作情况:A和B分别为WB检测细胞裂解液中Cx31、Cx26的表达情况;C为CO-IP检测Cx26突变体与Cx的互相作用
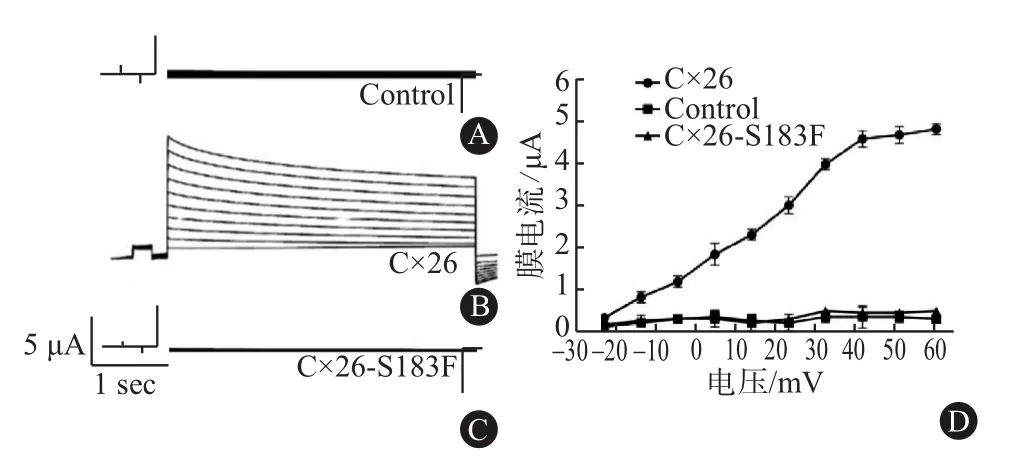
图4 膜片钳实验检测长岛型掌跖角化症(NPPK)野生型Cx26和突变体的膜电流:A为检测注射水的对照组卵母细胞的电流,B为检测单独注射Cx26的卵母细胞的电流,C为检测单独注射Cx26突变体的卵母细胞的电流,D为半通道I-V曲线图
2.6 Cx26突变体能增强Cx31半通道 CO-IP实验证明Cx31与异型半通道中的Cx26突变体结合,从而导致Cx26 NPPK突变体改变了Cx31的活性。因此验证了Cx26-S183F是否也会影响Cx31半通道活性。结果显示,单独注射Cx31的卵母细胞没有半通道活性(图5A),由Cx31和Cx26-S183F组成的异型半通道出现较高的电流(图5B)。以电流和电压作图,结果发现在电压的增加下,共表达Cx26-S183F和Cx31的细胞中的半通道活性显着增加,说明具有半通道功能。Cx31和Cx26-S183F共表达细胞产生的电流在-30、-20、-10、0、10、20、30、40、50、60 mV分别达到 0.135、0.21、0.235、0.3、0.315、0.38、0.40、0.41、0.44、0.46 μA,显著高于单独表达Cx31的细胞产生的电流(P=0.0316,图5C)。这些结果说明,当共表达形成异型连接子时,Cx31和Cx26突变体能够形成功能性半通道。
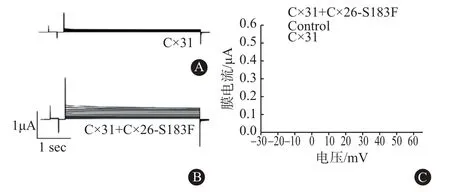
图5 膜片钳实验检测长岛型掌跖角化症(NPPK)Cx31和Cx26-S183F共表达时的膜电流:A为检测单独注射Cx31的卵母细胞的电流,B为检测注射Cx26-S183F+Cx31的卵母细胞的电流,C为半通道I-V曲线图
3 讨论
NPPK由丝氨酸蛋白酶抑制剂B7(SERPINB7)基因突变引起。在本实验中,测序发现2例病人的NPPK并不是由SERPINB7突变引起,通过测序发现一个与NPPK发生相关的GJB2突变。
当单独表达时,Cx26-S183F不能形成功能性同源半通道或间隙连接,但与Cx31共表达后,降低了Cx31间隙连接的活性,并增加了半通道活性。Cx31和Cx26突变体的免疫共沉淀显示形成异型连接子,Cx26突变体具有修饰Cx31半通道和缝隙连接的能力。
目前,大量研究表明间隙连接半通道在病理条件下能开放,为细胞间的物质交换提供通路。间隙连接半通道过量或长时间的开放会导致离子调节的失常,细胞内ATP浓度的变化和代谢产物丢失,最终导致细胞死亡[17]。不同间隙连接蛋白基因的突变可以导致同一种遗传学疾病,已有研究发现,Cx26突变与Cx43互作会引起掌跖角化病[18]。本项研究发现了Cx26-S183F与Cx31共表达会增强间隙连接半通道活性,从而引起NPPK。
Cx31在表皮上表达,而且Cx31突变会引起遗传性皮肤病,被称为变异性红皮肤角化病(EKV),其特征为躯干和四肢的皮肤过度角化,并伴有红斑出现[19]。研究发现,与Cx26突变相关的引起的皮肤病有KID综合征、PPK、先天性耳聋伴发角质厚皮病等[20-23]。目前为止关于Cx26突变引起皮肤病的机制有两种:(1)抑制其他角质形成细胞连接蛋白;(2)异常的半通道的形成。反式显性负效应作用会减少连接蛋白类型的数量从而影响表皮细胞间通讯,导致通道功能异常,而活跃的半通道会产生代谢物向细胞外释放,可能对邻近细胞产生不利影响,从而引发细胞凋亡[24]。
在Cx26和Cx31突变引起皮肤病的病人中,观察两者之间的临床特征存在相似性。四肢出现过度角化病变,这表明可能与NPPK出现的机制相似。除此之外,还有Cx26突变引起的Bart-Pumphrey综合征KHLS也有相似的临床症状,包括角化过度和白细胞减少症[25]。尽管症状相似,但仍需要进一步研究突变体确认其相关机制,Cx31-G8V突变的表达导致半通道的形成,允许钙离子流入细胞,从而造成KHLS[26]。有研究发现Cx30.3突变体会引起变异性红皮肤角化病(EKV),当Cx31和Cx30.3-F137L共同转染时,在细胞膜上检测到与Cx31形成间隙连接,并能形成异聚体连接子[27]。最近的一项研究表明,造成KID形成的Cx26-S17F突变体存在的情况下,Cx31半通道活性增加[28]。这与我们的研究结果一致,Cx26-S183F不能单独形成半通道或间隙连接,但在与Cx31共表达时可以增强半通道活性,从而引起NPPK。
这些研究结果表明Cx26在皮肤疾病中的重要性,并进一步突出了Cx26突变体和Cx31互相作用在遗传性皮肤病中的作用机制,为进一步治疗皮肤病提供一定的理论基础。
