五氯苯酚对淡水双壳类背角无齿蚌ρ型谷胱甘肽S-转移酶表达的影响
2020-01-09华春秀张科刘庆春李冰洁宋国英薛士鹏于瑞雪王中晓张庆远刘丽夏西超
华春秀,张科,刘庆春,李冰洁,宋国英,薛士鹏,于瑞雪,王中晓,张庆远,刘丽,夏西超,,*
1. 南阳医学高等专科学校基础医学部,南阳 473061 2. 平顶山学院医学院,平顶山 476000
五氯苯酚(PCP)广泛应用于农药、木材防腐剂和个人护肤品等工农业生产中,目前,在地表水、地下水、污水和饮用水中都不同程度地被检测到[1-3]。PCP存在能够给水生生物和人类生存带来显著的健康威胁,已被美国、中国和欧洲等国家和地区定为一种具有严重危害的持久性污染物[4-6]。在我国,PCP主要用于杀灭血吸虫的中间宿主,由于其持久性的存在,在大多数河流中仍然被广泛发现[1,7-8]。谷胱甘肽S-转移酶(GST)属于机体Ⅱ相生物转化系统酶类,催化谷胱甘肽(GSH)与广泛外源性或内源性有毒化合物亲电中心结合,如癌症化疗药物、化学致癌物、农药、除草剂和氧化应激产物[9-10]。在哺乳动物中,根据GST底物特异性、免疫学性质和蛋白质序列的同源性,把GST划分为α、μ、π、θ、σ、ζ和ω等7个大类[11]。在氧化应激条件下,GST表达水平被视为是一种适应性反应,GST表达被认为是受氧化应激动物的分子标志物[12-13]。背角无齿蚌(Anodontawoodiana)作为淡水底栖生物的重要类群,在淡水环境监测中具有重要作用[14-15]。前期研究发现,PCP处理对背角无齿蚌产生显著的毒性伤害和应激效应,具体机制有待进一步探讨[16]。在本研究中,从背角无齿蚌中克隆出ρ型谷胱甘肽S-转移酶完整基因序列并命名为ρ-GST,通过real-time PCR分析ρ-GST时空表达,为揭示PCP毒性效应奠定理论基础。
1 材料与方法(Materials and methods)
1.1 背角无齿蚌处理
背角无齿蚌购自南阳市水产市场,壳长(6.5±0.5) cm,处理之前,动物置于实验室自动水循环系统中适应养殖2周。PCP购自Sigma-Aldrich(Sigma-Aldrich, St. Louis, MO),溶解于二甲亚砜(DMSO)中以制备储备液。动物处理实验在长方形塑料盒(40 cm×25 cm;10 cm高)中进行,饲养采用人工模拟池塘水(每1 L去离子水中含48 mg NaHCO3、33 mg CaCl2·2H2O、60 mg MgSO4·7H2O和0.5 mg KCl)[16],每天更换水,动物喂食小球藻。为了确定ρ-GST组织分布,对来自同一塑料盒的5只动物进行解剖,取斧足、鳃、肝胰脏、闭壳肌、心脏、血淋巴和外套膜等组织。根据上述动物处理方法,将80只河蚌随机饲养于10个塑料盒中,每盒8只,分为对照组和PCP处理组,每组5个盒子。PCP处理组采用13.9 mg·L-1的PCP进行处理,对照组用同体积DMSO处理,水中DMSO浓度不超过3‰。0、1、3、6、9、12和15 d从每组中取出5只河蚌,解剖肝胰脏、鳃和血淋巴,液氮速冻,于-80 ℃保存。
1.2 总RNA提取和cDNA第一链的合成
总RNA提取采用TRIzol试剂(宝生物生物有限公司,大连),1.2%琼脂糖凝胶电泳检测RNA质量,具有完整rRNA条带的RNA用于合成cDNA,M-MLV试剂盒合成第一链cDNA,用作PCR反应模板。
1.3 背角无齿蚌ρ-GST核心片段的扩增
简并引物ρ-GST1和ρ-GST2分离ρ-GST cDNA保守区域片段,PCR产物连接至pMDT-19载体,双向测序。确定ρ-GST部分cDNA序列后,根据部分cDNA序列设计的特异性引物(表1),按照试剂盒要求,扩增ρ-GST cDNA 5’和3’区域序列,5’RACE和3’RACE的PCR产物进行测序和拼接。
1.4 序列和系统发育分析
分析ρ-GST序列,通过GenBank数据库搜索(www.ncbi.nlm.nih.gov/blast)进行BLAST程序比对;根据http://www.cbs.dtu.dk/services/SignalP预测信号肽;采用Simple Modular Architecture Research Tool (http://smart.embl-heidelberg.de/)预测蛋白质结构域;使用DANMEN分析程序对ρ-GST基因进行多序列比对;通过Swiss-model (http://swissmodel.expasy.org/)预测ρ-GST的蛋白质三维结构;使用MEGA5.0软件,采用临近法构建系统进化树。
1.5 ρ-GST mRNA水平定量检测
为了确定ρ-GST转录水平,采用SYBR Premix Ex TaqTM试剂盒并按照要求进行定量分析。β-actin作为内参基因,根据ρ-GST-F和ρ-GST-R引物常用PCR仪中分离靶基因(表1),琼脂糖凝胶电泳仅检测出一个条带,PCR产物测序,序列鉴别。使用ABI7500实时检测系统(Applied Biosystems,美国)进行real-time PCR,构建标准曲线,通过2-△△CT分析ρ-GST表达水平。
1.6 统计学处理
2 结果(Results)
2.1 背角无齿蚌ρ-GST分子结构
背角无齿蚌ρ-GST cDNA全长为1 026 bp,包含了57 bp的5’非编码区、291 bp的3’非编码区和一个678 bp的开放阅读框。开放阅读框为226个氨基酸组成的多肽,分子质量为26.80 kDa,等电点为6.38。终止信号(AATAAA)位于3’UTR的986~991处(图1)。预测氨基酸序列包括N端G-位(残基8~85)和C端H-位点(残基92~204)2个保守区域(图1)。
背角无齿蚌ρ-GST和其他水生软体动物GSTs之间的多重序列比对结果表明,所有GSTs的N-端区域同源性较高,并且相对保守,而C-端相对多变(图2)。在ρ-GST中,N端部分是谷胱甘肽S-转移酶催化的必要条件,通过稳定二硫键来激活被束缚的GSH。在该区域中,Try7、Ser13、Pro15、Phe38、His42、Lys43、Glu69和Ser70都是所有谷胱甘肽S-转移酶序列中的保守氨基酸,G-位点上这些氨基酸与GST结合有关(图2)。

图2 背角无齿蚌ρ-GST与其他物种GST序列多重比对注:G-位点保守氨基酸残基用星号表示。CgGST为长牡蛎谷胱甘肽S-转移酶(EKC36795.1),HdGST为皱纹盘鲍(Haliotis discus discus) 谷胱甘肽S-转移酶A(ABO26605.1),LeGST为南极帽贝(Laternula elliptica)ρ型谷胱甘肽S-转移酶(ACM44933.1),RpGST为菲律宾蛤仔(Ruditapes philippinarum)ρ型谷胱甘肽S-转移酶(AEW46331.1)。Fig. 2 Multiple alignment of ρ-GST of Anodonta woodiana compared with other GSTSsNote: CgGST is Crassostrea gigas glutathione S-transferase A (EKC36795.1); HdGST is Haliotis discus discus glutathione S-transferase A (ABO26605.1); LeGST is Laternula elliptica rho-class glutathione S-transferase (ACM44933.1); RpGST is Ruditapes philippinarum rho-class glutathione S-transferase (AEW46331.1). The conserved residues of G-sites are marked with asterisk.
2.2 AwGST1和AwGST2的二级和三级结构分析
ρ-GST蛋白质二级结构有12个α-螺旋和4个β-折叠组成(图3A)。在N端G-位点有3个α-螺旋和4个β-折叠,C端的H-位点包含8个α-螺旋。ρ-GST的三维结构与其他物种ρ型谷胱甘肽S-转移酶有高度的相似性(图3B)。
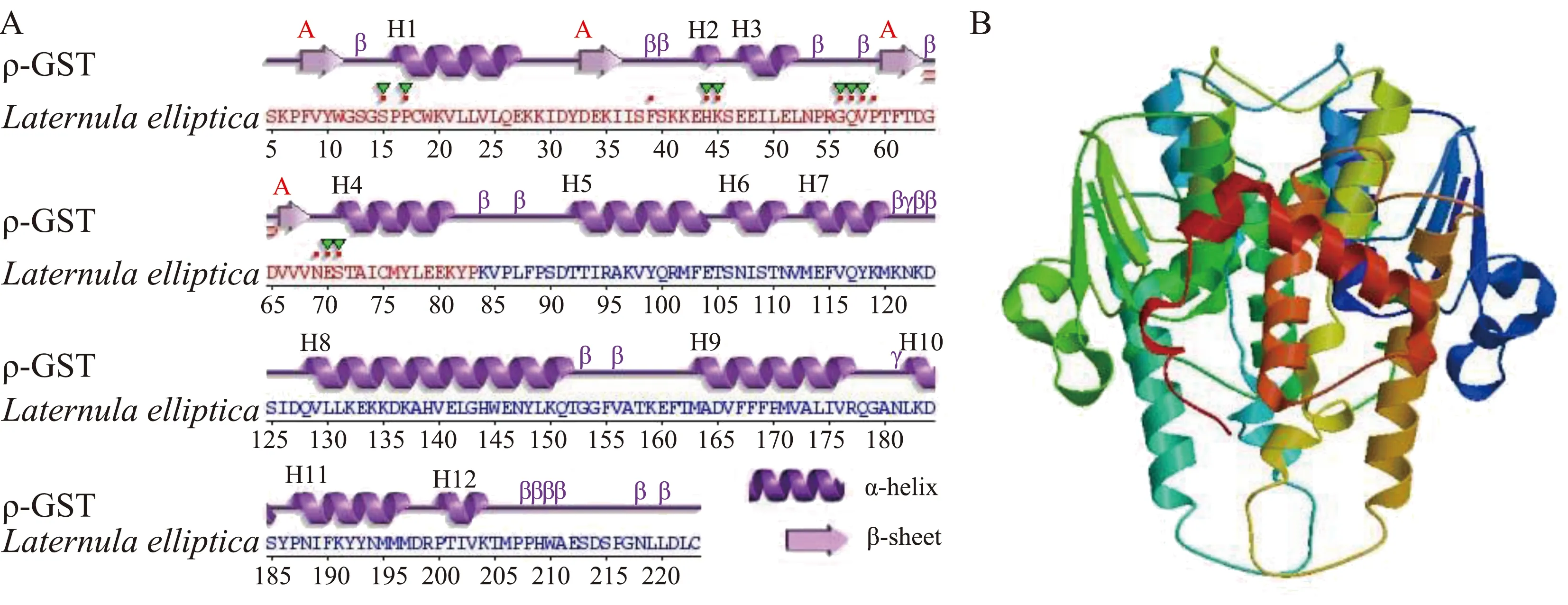
图3 背角无齿蚌ρ-GST二级和3D结构预测注:A. ρ-GST二级结构;B. ρ-GST的3D结构。Fig. 3 Predicted secondary and 3D structures of ρ-GST deduced amino acids of Anodonta woodianaNote: A. The secondary structure of ρ-GST; B. The 3D structure of ρ-GST.
2.3 背角无齿蚌ρ-GST系统进化
背角无齿蚌ρ-GST氨基酸序列与腹足纲皱纹盘鲍GST有55%同源性,与双壳纲长牡蛎(Crassostreagigas)GST有55%同源性,与腹足纲南极帽贝ρ-GST有50%同源性,与菲律宾蛤仔GST有46%同源性(表2)。
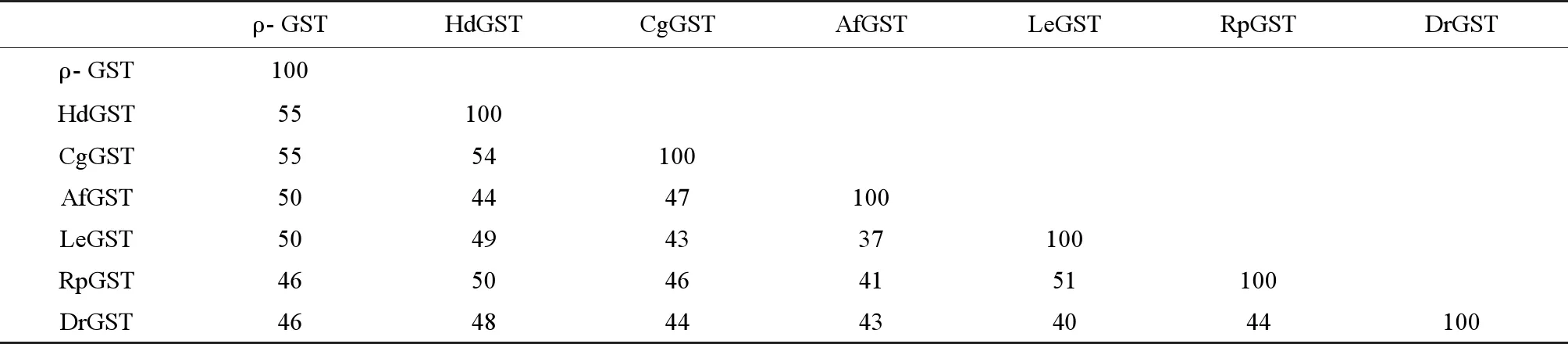
表2 背角无齿蚌ρ-GST与其他物种GST序列同源性比较Table 2 Percentage of identity respectively for pairwise alignment of ρ-GST protein sequences between Anodonta woodiana and other species
注:HdGST为皱纹盘鲍谷胱甘肽S-转移酶A(ABO26605.1),CgGST为长牡蛎谷胱甘肽S-转移酶(EKC36795.1),AfGST为栉孔扇贝(Azumapectenfarreri)ρ型谷胱甘肽S-转移酶(ACF25903.1),LeGST为南极帽贝ρ型谷胱甘肽S-转移酶(ACM44933.1),RpGST为菲律宾蛤仔ρ型谷胱甘肽S-转移酶(AEW46331.1),DrGST为斑马鱼(Daniorerio)ρ型谷胱甘肽S-转移酶(NP_001038525.1)。
Note: HdGST isHaliotisdiscusdiscusglutathione S-transferase A (ABO26605.1); CgGST isCrassostreagigasglutathione S-transferase A (EKC36795.1); AfGST isAzumapectenfarrerirho-class glutathione S-transferase (ACF25903.1); LeGST isLaternulaellipticarho-class glutathione S-transferase (ACM44933.1); RpGST isRuditapesphilippinarumrho-class glutathione S-transferase (AEW46331.1); DrGST isDanioreriorho-class glutathione S-transferase (NP_001038525.1).
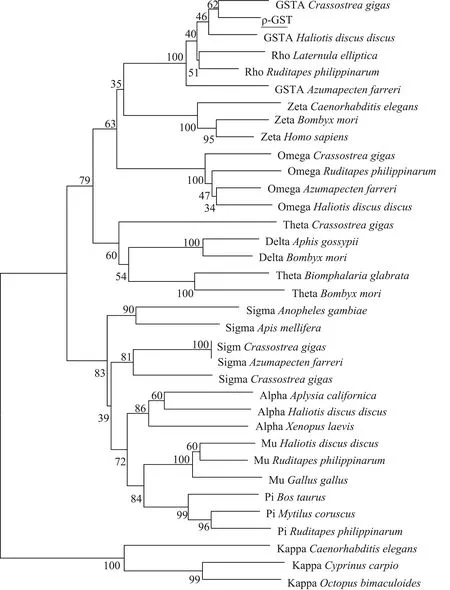
图4 背角无齿蚌ρ-GST氨基酸序列使用邻接法构建的系统进化树Fig. 4 Phylogenetic relationship of ρ-GST of Anodonta woodiana according to neighborhood-joining method
系统进化树结果显示,背角无齿蚌ρ-GST与软体动物谷胱甘肽S-转移酶A类群和ρ型GST亲缘关系较近,谷胱甘肽S-转移酶A类群和ρ型GST分为了一组(图4)。研究结果显示,从选择的物种中,同一类型GST的不同个体之间保持较高的bootstrap值,特别是在θ型GST中,昆虫家蚕(Bombyxmori)和脊椎动物之间bootstrap值达到99%。值得注意的是,2种类型GST之间,bootstrap值明显减小,如ω型GST和ζ型GST、ω型GST和δ型GST、ζ型GST和σ型GST等。
2.4 背角无齿蚌ρ-GST表达水平的组织分布
背角无齿蚌ρ-GST广泛表达于斧足、外套膜、闭壳肌、心脏、肝胰腺、血淋巴和鳃(图5)。背角无齿蚌ρ-GST在肝胰腺中表达水平最高,在鳃、血淋巴、心脏、斧足和闭壳肌中次之,外套膜中表达水平较低(图5)。
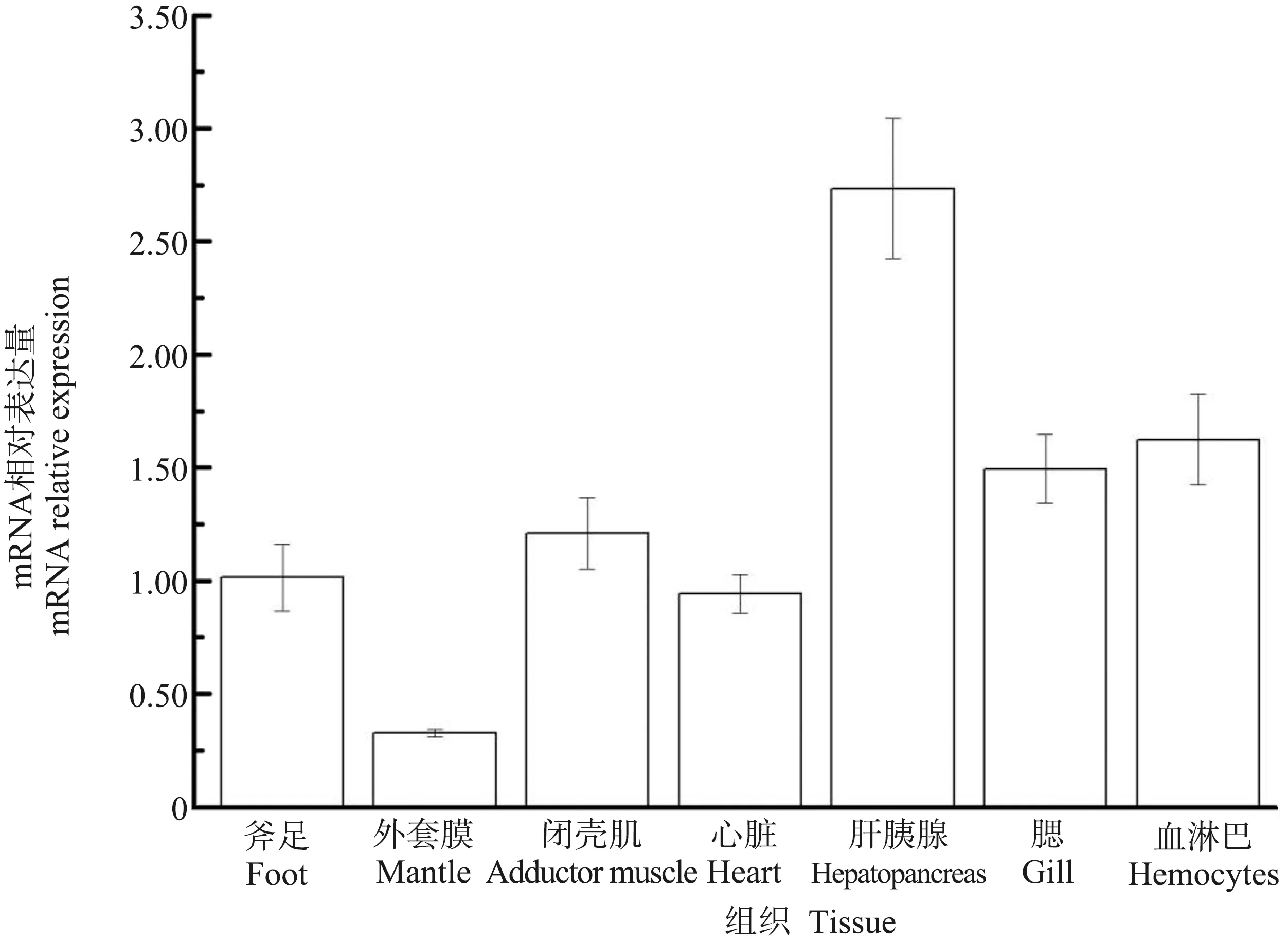
图5 背角无齿蚌ρ-GST基因的空间表达注:每组数据来源于5只动物。Fig. 5 Real-time PCR analysis of ρ-GST transcript from different tissues of Anodonta woodianaNote: n = 5 replicates.
2.5 PCP对背角无齿蚌ρ-GST表达的影响
在肝胰腺中,与第0天相比,背角无齿蚌ρ-GST表达在第1~15天出现了明显的波动。PCP处理对背角无齿蚌ρ-GST表达具有显著影响。PCP处理后背角无齿蚌ρ-GST mRNA水平呈现时间依赖的上调模式。与对照组相比,ρ-GST表达水平在第1天、第3天和第15天分别增加18.18%、82.88%(P<0.05)和2.43倍(P<0.01) (图6)。与对照组相比,PCP处理后背角无齿蚌鳃中ρ-GST表达水平增加了1.44倍以上(P<0.05) (图7)。与对照组相比,PCP处理后背角无齿蚌血淋巴中ρ-GST表达水平在第3至15天显著升高(图8)。
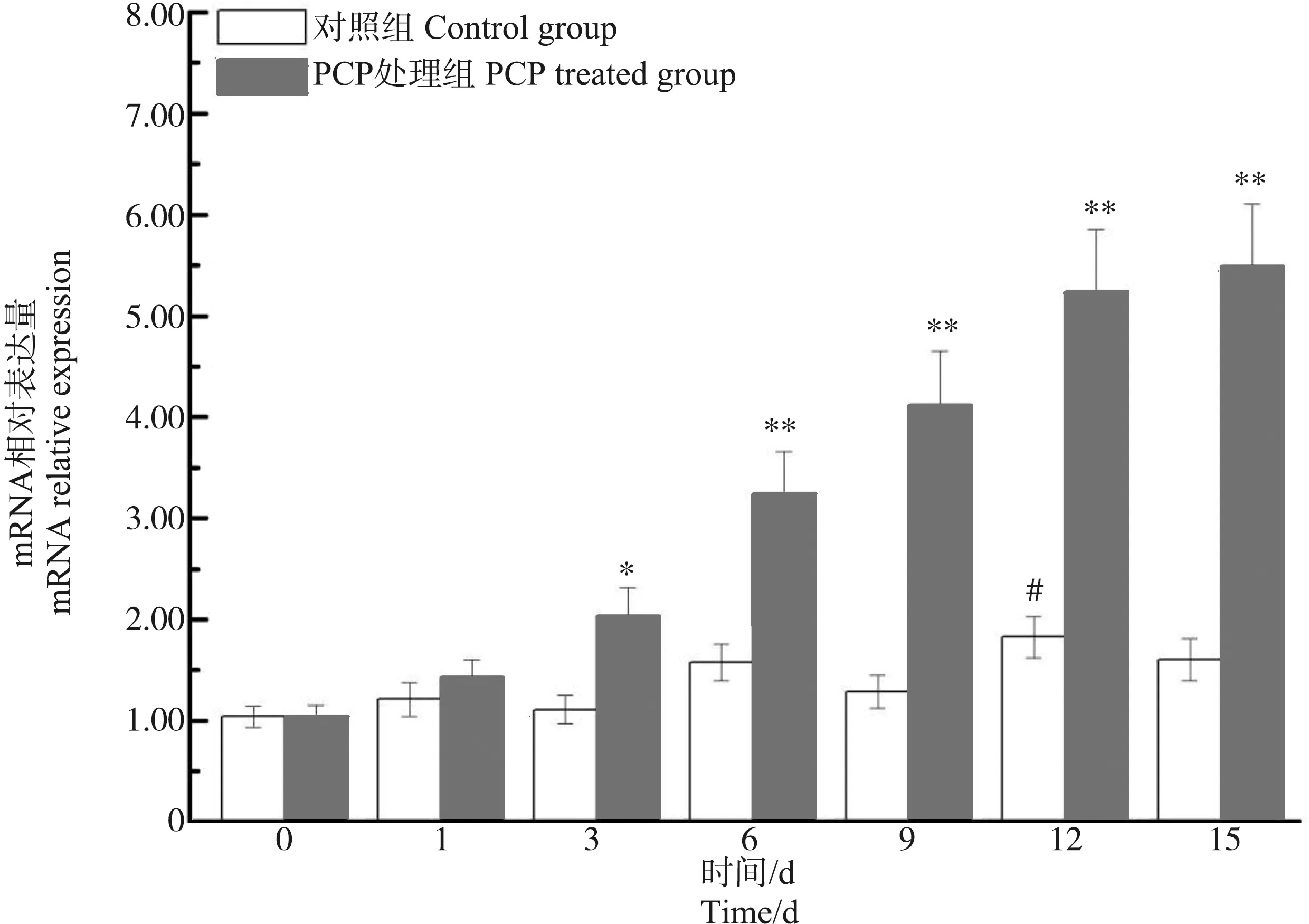
图6 五氯苯酚(PCP)对背角无齿蚌肝胰腺ρ-GST基因表达的影响注: n=5/组/时间点;*、**表示与相应对照组相比,有显著差异(P<0.05、P<0.01);#表示与0 d相比,有显著差异(P<0.05)。Fig. 6 Temporal expression of ρ-GST in hepatopancreas of Anodonta woodiana after pentachlorophenol (PCP) challenge as measured by quantitative real-time RT-PCRNote: Bars represent means ± SE; n=5/each group/each time point. * P<0.05, ** P<0.01 vs control group at the same time. # P<0.05, ## P<0.01 vs control group at the day 0.
3 讨论(Discussion)
ρ-GST有N端区域和C-端区域2个不同区域;多重序列比对表明,ρ-GST的N端G-位点相对保守,而C端多变,提示这些特征与GST的功能有关。一般来说,供GSH结合的G位点具有高度特异性包含11个高度保守氨基酸:Tyr7、Arg13、Trp38、Lys46、Gln53、Leu54、Pro55、Gln66、Ser67、Glu98和Asp99[17-18]。前期研究表明,Tyr在稳定结合GSH方面发挥重要作用[17-18],提示ρ-GST的N端保守氨基酸残基有助于结合GSH。H-位点是亲电底物的结合点,C端的H-位点相对多变与适应更广泛底物的结构有关[19-20]。
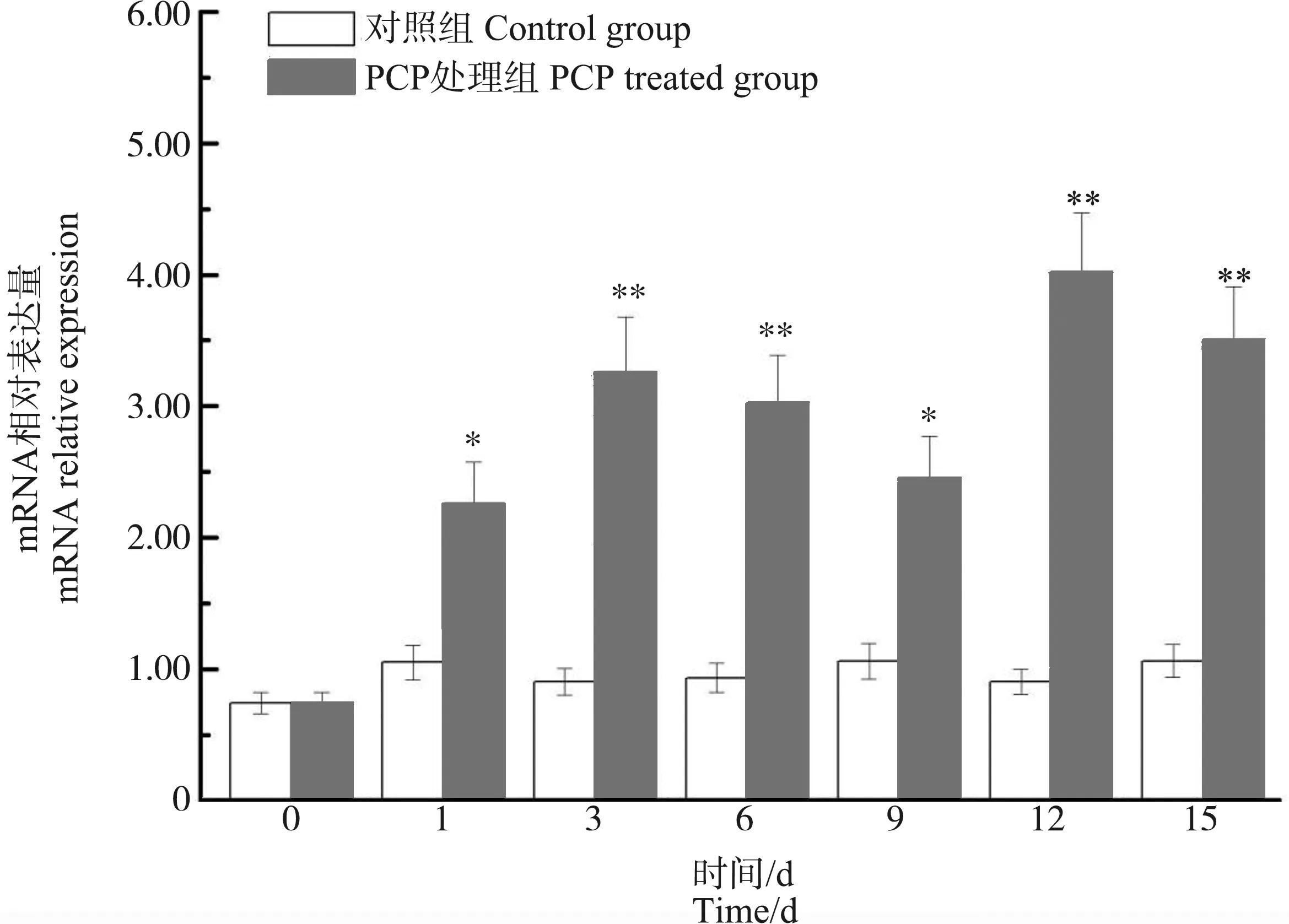
图7 PCP对背角无齿蚌鳃ρ-GST基因表达的影响注:n=5/组/时间点;*、**表示与相应对照组相比,有显著差异(P<0.05、P<0.01)。Fig. 7 Temporal expression of ρ-GST in gill of Anodonta woodiana after PCP challenge as measured by quantitative real-time RT-PCRNote: Bars represent means ± SE; n=5/each group/each time point. * P<0.05, ** P<0.01 vs control group at the same time.
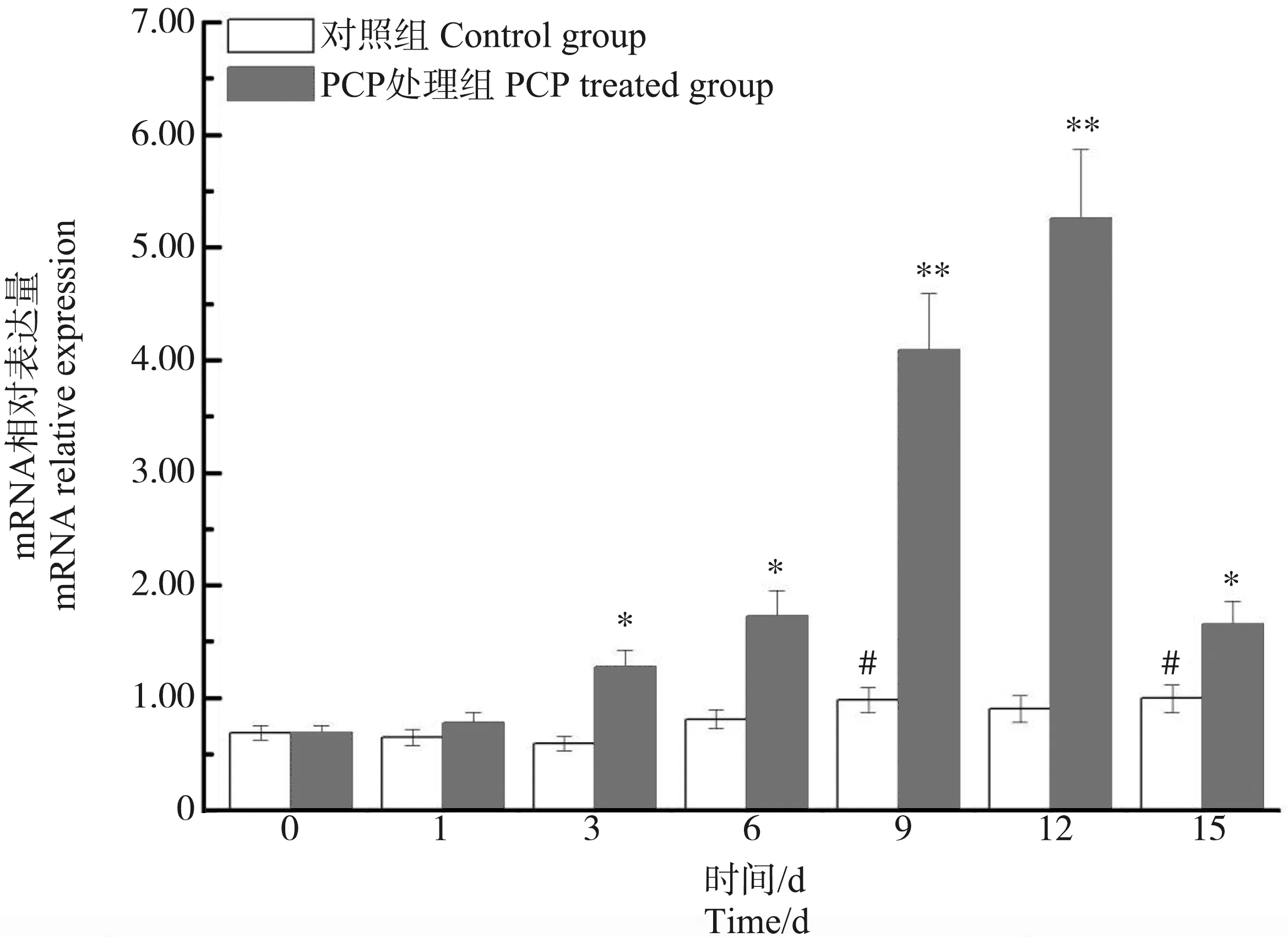
图8 PCP对背角无齿蚌血淋巴ρ-GST基因表达的影响注:n=5/组/时间点;*、**表示与相应对照组相比,有显著差异(P<0.05、P<0.01);#表示与0 d相比,有显著差异(P<0.05)。Fig. 8 Temporal expression of ρ-GST in hemocytes of Anodonta woodiana after PCP challenge as measured by quantitative real-time RT-PCRNote: Bars represent means ± SE; n=5/each group/each time point. * P<0.05, ** P<0.01 vs control group at the same time. # P<0.05 vs control group at the day 0.
多序列比对结果显示,ρ-GST氨基酸序列与长牡蛎和腹足纲皱纹盘鲍GST-A序列很相似,并与ρ型GST具有高度同源性。基于系统进化分析数据和氨基酸序列比对结果,ρ-GST应归为ρ型GST。研究已表明,序列相似性程度在GST分类中没有明确标准,GST同工酶的演化关系仍然存在广泛争议[21]。大家普遍认为,在一类GST个体中同源性超过40%划分为一个类群,同源性小于25%类群划分为其他类群[21]。θ类GST第一个关于子家族演化的描述表明,θ类GST在从有氧细菌到更高真核生物中都存在,呈现广泛分布,序列具有高度保守性,是较为原始的类群[22]。σ类GST是从哺乳动物α、μ和π这3个类群中分离出来的,ρ和θ从α、μ、ρ和θ中分离出来[23]。由此可见,GST类群进化经历了从无脊椎动物到脊椎动物的复杂里程。
背角无齿蚌ρ-GST在肝胰腺中表达水平最高,在鳃、血淋巴、心脏、斧足和闭壳肌内中等表达,在外套膜表达水平较低,这提示ρ-GST这种表达模式可能与清除活性氧以及维持氧化和抗氧化细胞之间平衡有关。正常情况下,活性氧簇(ROS)保持相对较低水平,ROS会被一系列抗氧化剂酶迅速消除,以维持ROS水平和抗氧化剂酶活性之间的均衡。在双壳类动物,肝胰腺是重要代谢器官,在解毒和外源性物质的降解方面发挥重要的作用[24]。软体动物GST能降解水体农药、有机污染物和其他有毒化学物质[25]。因此,肝胰腺ρ-GST mRNA水平高表达与环境中出现外源性有毒物质降解有关。
在整个实验过程中,PCP明显诱导背角无齿蚌肝胰腺、鳃和血淋巴中ρ-GST的表达,提示ρ-GST表达水平上调有助于参与PCP解毒过程和抗氧化过程。对于机体而言,ROS主要包括过氧化物阴离子、过氧化氢、烷基过氧化物、单线态氧和羟基自由基[26]。背角无齿蚌是滤食性淡水动物,PCP易沉积和过量积累在其体内,可以催化生成ROS[3-4]。GST广泛分布在所有活细胞中并参与机体有毒物质降解,GST通过与毒素结合,能够主动或者被动地与致癌物、治疗制剂和氧化应激产物等外源性/内源性有毒物质结合[27-28],促进细胞排毒和自我保护的作用。研究表明,正常情况下GST持续表达与机体解毒密切相关,而GST的高表达参与保护细胞免受内源性氧化应激的影响[27-29]。已有研究表明,诸多外界因素诱导GST表达,GST被认为是一种对抗异种生物化学毒性的有效解毒酶。高水平GST与农药降解密切有关[30-31]。褐飞虱(Nilaparvatalugens)GST水平增高有助于抵抗氧化损伤,保护组织[32]。综上所述,背角无齿蚌中ρ-GST表达上调在降低PCP导致的氧化应激和细胞损伤过程中发挥着积极作用。
PCP处理后ρ-GST表达水平在不同组织表现出不同特征。对于该现象,一种解释可能与这些组织不同生理功能有关。软体动物肝胰腺具有肝和胰腺双重功能,参与消化和中和大量有毒物质[33-34]。鳃位于壳腔内,通过水体流动实现呼吸功能,直接与外部环境接触,上皮组织非常薄[35],鳃也是环境污染物作用的重要靶器官之一[36]。双壳类血淋巴是防御病原侵染的重要防线,也是减少ROS生成的重要防线,通过吞噬作用和呼吸爆发过程消除环境污染物[37]。众所周知,呼吸爆发过程与双壳类血淋巴中重金属和污染物所产生的氧化应激密切相关[37-38]。呼吸爆发在消除重金属和污染物产生的毒性方面起着潜在的作用,一些复杂因素可能参与肝胰腺、鳃和血淋巴中ρ-GST表达调控。另一种解释是ρ-GST可能在不同的组织和不同的发育阶段扮演不同的角色。
与脊椎动物相比,软体动物主要依靠自身的免疫能力,而血淋巴细胞是先天防御措施的主要途径之一。维持机体稳态是一个复杂过程,需要多个器官参与。在环境压力下,一系列抗氧化酶表达在其中发挥重要的作用,高表达涉及保护细胞,抵抗内源性氧化应激。背角无齿蚌ρ-GST在肝胰腺、鳃和血淋巴中表达显著增加有助于清除PCP处理产生的ROS,提高细胞氧化应激能力,实现机体稳态。
