河-库连续体中溶解性无机碳及其同位素的时空分异特征*
——以澜沧江云南段为例
2020-01-09郭梦京周孝德谢曙光
袁 博,吴 巍,郭梦京,周孝德**,谢曙光
(1:西安理工大学西北旱区生态水利国家重点实验室,西安 710048) (2:北京大学环境科学与工程学院,北京 100871)
河流是联系陆地与海洋生态系统的主要纽带,水体中携带的溶解性有机质(DOM)的迁移转化是维持不同尺度下地表水营养盐平衡、碳平衡、沉积物平衡的关键组成部分,并在很大程度上控制着水生态系统的功能[1]. 河-库生态系统是碳(C)、氮(N)、磷(P)等生源物质的重要“源汇”转化场所[2];碳作为所有水生生物必需的生源要素,也是水生生物地球化学研究中最为关注的元素之一[3]. 溶解性无机碳(DIC)作为河流及水库等淡水系统中最丰富的无机形态碳[4],是全球碳循环与大气、陆地和海洋之间碳相互作用的重要组成部分[5]. DIC与溶解性有机碳(DOC)之间耦合关系与相互转化在影响水体中光合作用及呼吸作用、有机质的氧化分解、水体养分的生物有效性以及作为微生物底物重要来源等方面发挥着重要作用[6]. 在阐明碳在大气-陆地-海洋系统中的循环机制方面,之前的研究已经认识到在河流等地表水输移过程中确定DIC来源和归宿的重要性[7-8]. 通过水体中DIC(δ13CDIC)和DOC(δ13CDOC)浓度和碳同位素的组成可以确定这些体系中溶解性碳的种类和浓度[9].
湖泊与水库等通常是作为单独的河流溶解性碳输移实体来进行研究的,而针对河流-水库/湖泊等连续体中溶解性碳的输移机制仍然缺乏深入探索[4]. 河流是单一的水流传输通道或是包括河漫滩在内的一个更大的河网系统[10]. 因此,在不考虑河网内有湖泊或水库存在的情况下,河流中碳的输移主要按照从源头到河口的连续统进行研究[11-12]. 然而,河流和水库形成的组合体应被视为水-陆两相耦合的碳输移通道和反应系统[13]. 因此,应将河网中水库对碳的滞留及转化过程涵盖在内,以量化河流-水库连续体的生物地球化学作用. 当前国内外大多数对溶解性碳动力学的研究都集中在寒带和温带地区[14-15],其结论可能并不一定反映出热带及亚热带流域的研究结果[14],且大部分研究仍集中在单一湖泊/水库上,鲜有针对大型河流(流域面积>200 km2)上的梯级水库DIC来源和时空变化特征的研究.
在水利工程梯级开发引发的河-库连通性阻隔的背景下,针对河-库系统生物地球化学循环的研究越来越受关注,这已经成为水利、环境及地球科学新兴的交叉研究前沿. 梯级水库中DIC的生物地球化学循环与单级水库/湖泊在影响因素和累积效应上存在显著差异,对其生物地球化学过程的认识还很有限[16]. 澜沧江-湄公河作为亚洲最大国际河流,其水资源利用和生态环境影响备受下游国家关注[17],而澜沧江云南段作为中国西南地区河流水电开发的重要基地,其干流集中了若干不同建成时期、不同污染来源、不同调节方式的水库,但对其DIC来源及归趋特征的研究鲜有报道. 基于此,本研究以西南地区梯级开发重点河流——澜沧江云南段河流-梯级水库连续体为研究对象,通过识别DIC在水体中的浓度输移变化特征和δ13CDIC同位素组成,确定控制DIC动力学的主要来源、输移转化的地球化学过程及驱动因素,探讨在不同水文时期水库在河-库连续体中溶解性碳输移的源汇机制,对维护河流生态健康和促进流域生态环境管理具有重要研究价值.
1 研究方法及材料
1.1 研究区概况
澜沧江-湄公河发源于青藏高原唐古拉山的格尔吉河和鄂穆楚河,两河自青海入西藏昌都汇合后成澜沧江,干流全长约4500 km,在中国境内全长2179 km,流域面积约164800 km2,多年平均径流量475 km3,天然落差约4583 m. 澜沧江云南段自云南省维西县入省境,于勐腊县出境入老挝,全长1240 km,区间流域面积90200 km2,落差1780 m,平均比降1.45‰. 澜沧江云南段主要地形特点为多高山峡谷,沟谷多呈V字形,随着河流深切谷底,高差悬殊,峰谷相对高差超过1000 m. 下游滇西南地区丘陵和盆地交错,属于亚热带或热带季风气候,干(11月-次年4月)、湿(5-10月)两季分明,约85%的降水量集中在湿季[18]. 自上而下较大的支流有沘江、黑惠江(漾濞江)、罗扎河、小黑河、小黑江(威远江)、黑河(扎糯江)、罗梭江(补远江)、南腊河等. 根据澜沧江梯级水电开发规划将云南段分为上游、中游和下游段,上游段长约489 km,天然落差1036 m,规划有1库7级水电站,自上而下依次为古水、乌弄龙、里底、托巴、黄登、大华桥和苗尾,总装机容量为9580 MW. 中游段有1库4级水电站,自上而下分别是功果桥、小湾、漫湾和大朝山,总装机容量8110 MW. 下游段按照1库4级规划,自上而下分别为糯扎渡、景洪、橄榄坝和勐松,勐松电站目前开发条件不太确定,其余3个电站合计装机容量8360 MW,截止2018年已建成7级水电站,已建成水电站主要信息见表1.

表1 澜沧江云南段已建梯级水电站主要指标
1.2 样品采集及预处理
本研究共计采样2次,分别于2018年8月11日-25日和2019年1月14日-25日从澜沧江入云南省界沿程向下游采样(图1). 澜沧江云南段上游规划河段比降较大,水流湍急,该河段未开设航道,在无船只的情况下,从桥梁或岸边以及尽可能远离河岸线的地方采集样品,该河段设置9个采样断面,分别为盐井(YJ)、古水(GS)、云岭(YL)、乌弄龙(WNL)、里底(LD)、白济汛(BJX)、托巴(TB)、黄登(HD)和大华桥(DHQ). 下游梯级水库河段采用走航船进行采样,根据水库形态结构和支流分布情况,从库尾到坝前将各水库依次划分为河流区、过渡区和湖泊区,按照水库回水长度和现场采样情况在每个分区设置1~2个采样断面;同时考虑支流的影响,分别在功果桥水库沘江(BJ)、小湾水库黑惠江(HHJ)和糯扎渡水库威远江(WYJ)等有大型支流汇入的主库区回水以上约1.5 km处设置采样断面. 其中,苗尾库区设置3个采样断面(MiaoW1~MiaoW3),功果桥库区及支流设置5个采样断面(GGQ1~GGQ4,BJ),小湾库区及支流设置8个采样断面(XW1~XW7,HHJ),漫湾库区设置8个采样断面(MW1~MW8),大朝山库区及支流设置5个采样断面(DCS1~DCS5),糯扎渡库区及支流设置10个采样断面(NZD1~NZD9,WYJ),景洪水库设置5个采样断面(JH1~JH5),景洪水库下游橄榄坝(GLB)和勐腊(ML)分别设置1个采样断面. 每个采样断面分别在距离河流左、右岸约10 m及河道中心布设采样点,将断面左、中、右所采水样混合后作为断面代表水样,每个断面采集3瓶混合水样作为平行样待测.
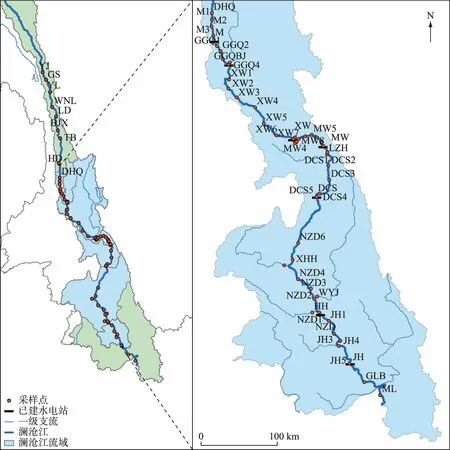
图1 澜沧江云南段研究区及采样点(盐井(YJ)、古水(GS)、云岭(YL)、乌弄龙(WNL)、里底(LD)、白济汛(BJX)、托巴(TB)、黄登(HD)和大华桥(DHQ))Fig.1 Study area and sampling sites in the Yunnan Section of Lancang River
现场采用便携式有机玻璃采水器采集水面以下0.5 m水样,用450℃预灼烧Whatman GF/F微孔滤膜过滤200 mL滤液于棕色聚乙烯瓶中保存,聚乙烯瓶采样前冲洗3次,加饱和HgCl2抑制微生物活性,不留气泡,采用Parafilm封口膜密封保存于4℃车载冰箱,用于δ13CDIC测定. 另取200 mL滤液装入预酸洗的玻璃瓶中,密封冷藏,所有样品分析测试均在采样后两周内完成.
1.3 水体理化指标及δ13C同位素分析

溶解性无机碳同位素(δ13CDIC)采用Atekwana等[19]的方法测定,水样带回实验室后注入已抽真空并放有浓磷酸和小磁棒的玻璃瓶中,50℃水浴加热,采用高真空线萃取,冷阱分离,收集纯CO2气体再用气体稳定同位素质谱仪(MAT252/253)测定δ13CDIC. 测定的值以千分比为单位(‰ ),分析误差<±0.1‰. 溶解性有机碳同位素(δ13CDOC)采用熔封石英管高温燃烧法测定[19],取适量预处理样品(约含0.5~1.0 mg C)与2 g线装CuO混合装入石英管内,经真空线系统抽真空后熔封,于马弗炉内850℃下燃烧5 h,缓慢待冷却后,导出CO2,经纯化系统纯化后使用气体稳定同位素质谱仪(MAT252/253)测定δ13CDOC. 有机质的δ13C对应国际标准V-PDB(Vienna PeeDee Belemnite),以国际纤维素标样IAEA-C3(δ13C =-24.91‰)为参考标准,分析误差<±0.2‰. δ13CDIC与δ13CDOC的计算公式为:
δ13C(‰)=[(Rsample-Rstandard)/Rstandard]×103
(1)
式中,Rsample表示测量样品的C同位素比值,Rstandard表示国际标准C同位素比值.
2 结果
2.1 澜沧江梯级河段水化学特征
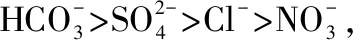
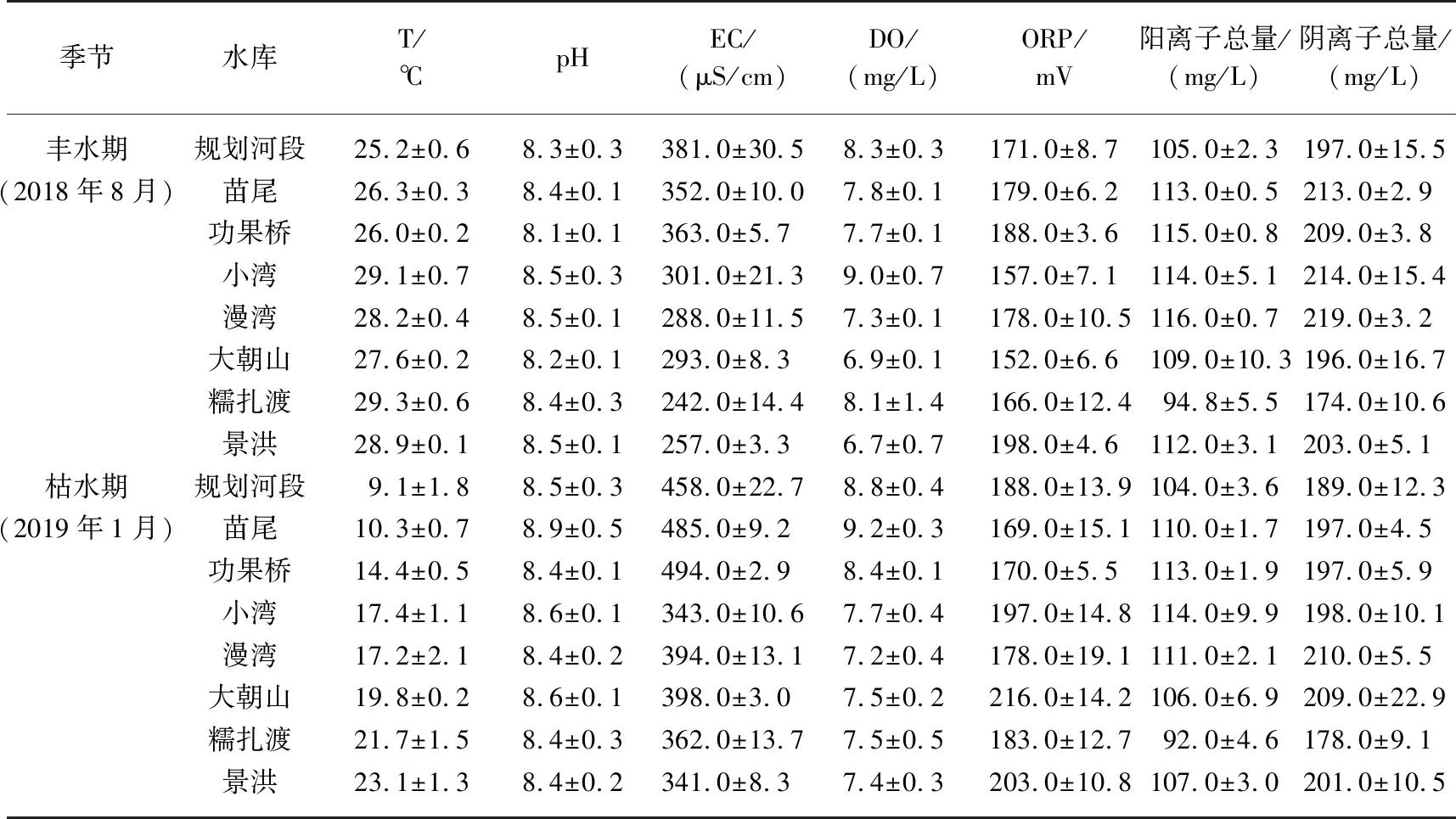
表2 研究河段及水库水体主要水化学参数
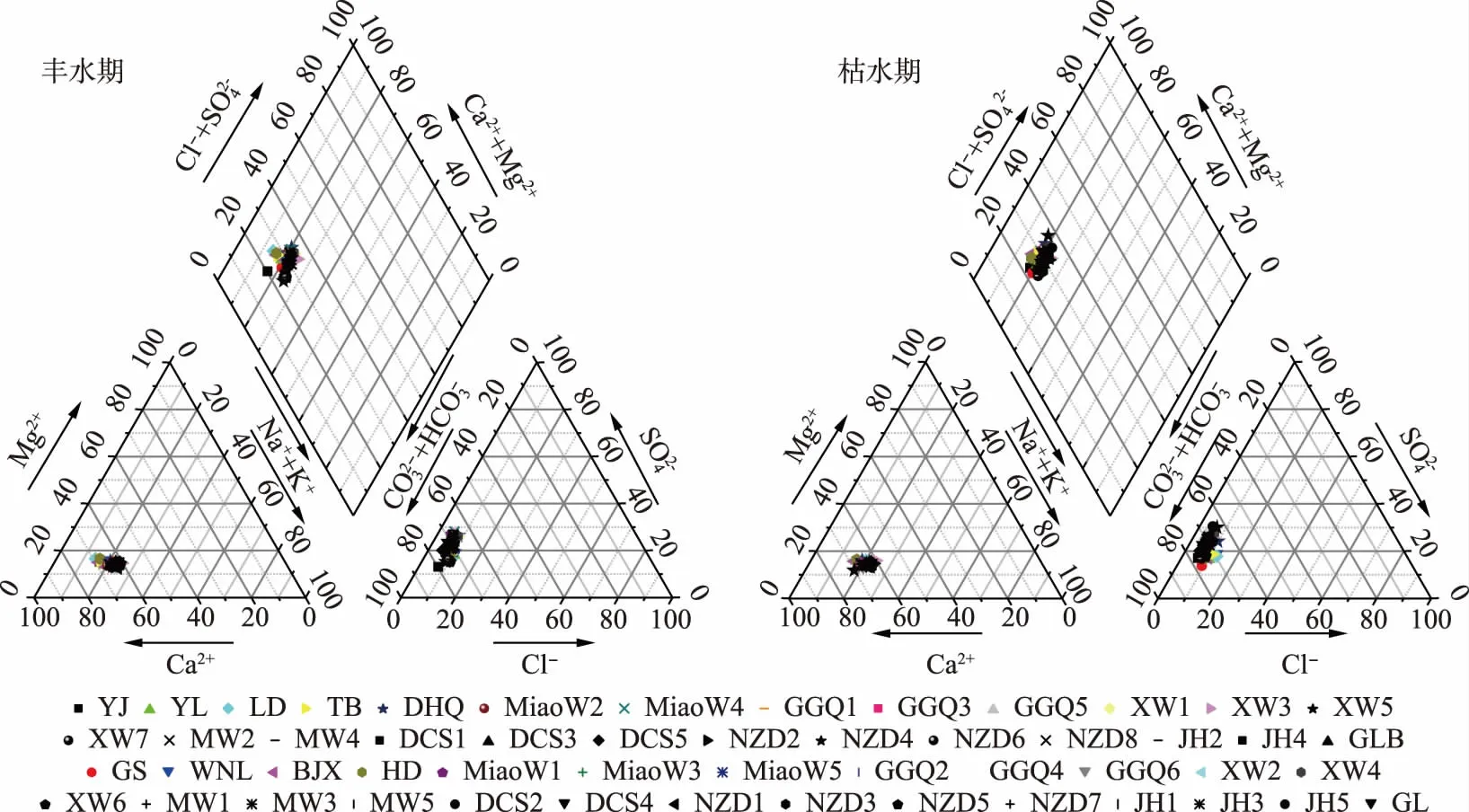
图2 澜沧江云南段水化学离子Piper图Fig.2 Piper plot of hydrochemical ions in Yunnan Section of Lancang River
2.2 澜沧江梯级河段DIC浓度及δ13CDIC组成的时空变化特征
在河流纵向梯度上,DIC浓度在丰水期和枯水期的变化趋势接近,从上游天然河段到下游梯级水库群均表现为先升高再降低再升高的趋势(图3a),丰水期的最高值出现在小湾库区河段,枯水期的最高值出现在功果桥水库河段,分别为3.38和4.05 mmol/L;丰水期和枯水期的最低值均出现在糯扎渡库区的上游河段,分别为1.54和2.36 mmol/L. 在时间变化上,澜沧江梯级河段DIC在丰水期(8月)的浓度为1.54~3.38 mmol/L,平均值为2.59±0.44 mmol/L;枯水期(1月)浓度为2.36~4.05 mmol/L,平均值为3.30±0.37 mmol/L,枯水期DIC浓度明显高于丰水期. 总体来看,梯级开发河段的DIC浓度在丰水期和枯水期均高于上游规划河段,澜沧江云南段δ13CDIC值在丰水期较枯水期偏负(图3b),介于-9.60~-7.70‰之间,平均值为-8.52‰±0.38‰;冬季枯水期则偏正,在-7.60‰~-5.70‰之间,平均值为-6.95‰±0.53‰,δ13CDIC同位素组成与DIC浓度变化表现出相似的特征,夏季丰水期澜沧江梯级水库河段中δ13CDIC值明显低于冬季枯水期. 在河流纵向梯度上,除两个运行时间较长梯级水库:漫湾(26年)和大朝山(18年)的δ13CDIC偏负外,其他河段及库龄较短的梯级水库整体变化比较平稳,澜沧江云南段δ13CDIC值整体呈现微弱升高的趋势. 随着水库运行时间的延长,δ13CDIC值呈现偏负趋势,表层水体可能体现最为明显.
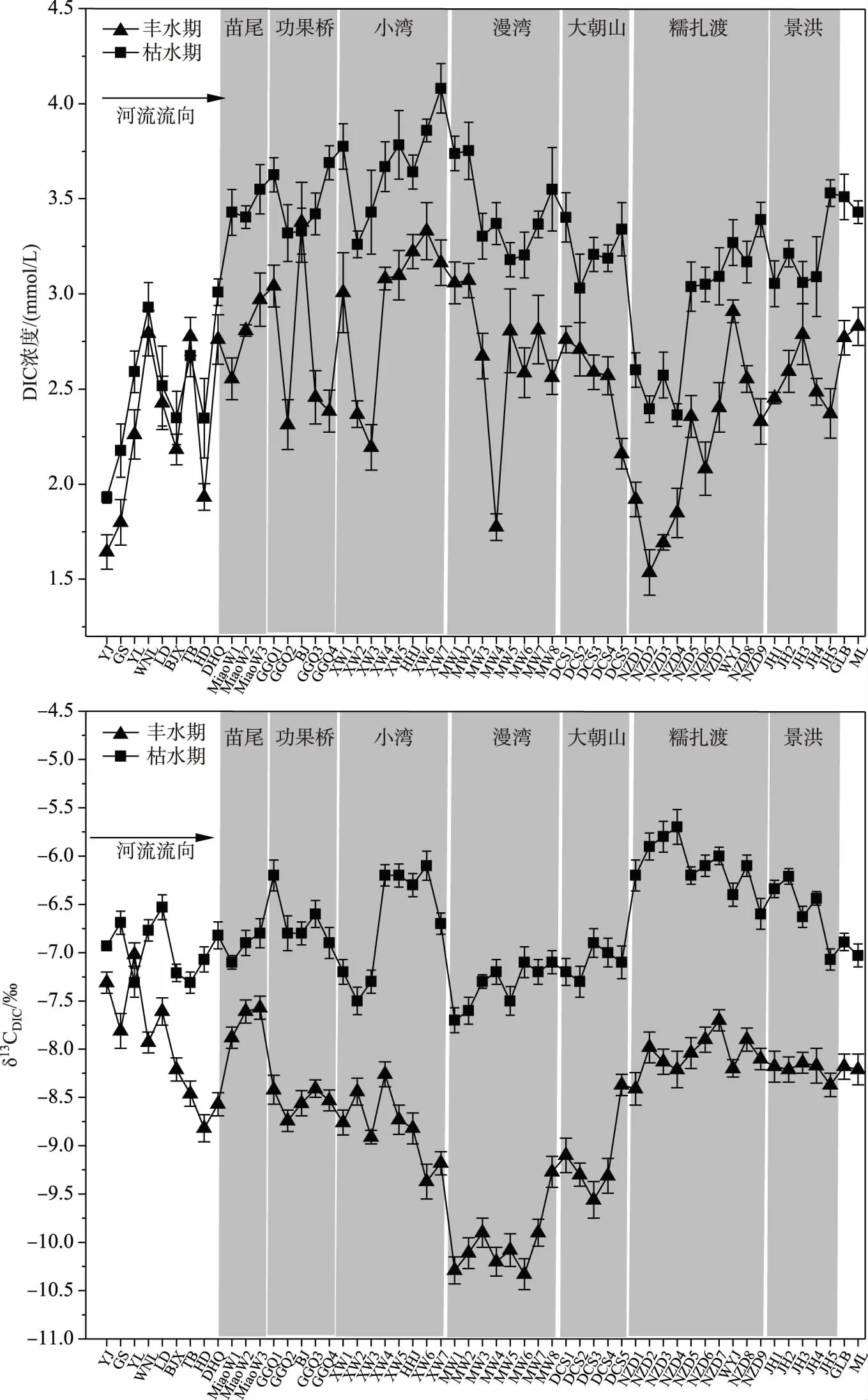
图3 澜沧江梯级河段水体中DIC浓度和δ13CDIC的时空变化Fig.3 Spatial and temporal variations of DIC concentration and δ13CDIC in the cascade reservoir reach of Lancang River
3 讨论
3.1 澜沧江梯级水库DIC及δ13CDIC的物源辨析
εaq-g=-(0.0049±0.003)T-(1.31±0.06)
(2)
(3)
(4)

3.2 梯级开发河段DIC浓度及δ13CDIC的变化特征
河流DIC浓度和δ13CDIC值的时空变化受多种因素影响,河流水电开发等人类扰动对其影响明显. 就单一水库、湖泊及天然河流的研究中,喻元秀等[30]对乌江上游新建的洪家渡水库中DIC的研究发现,入库水体中DIC浓度低于出库水体,出库水体中δ13CDIC值比入库水体和库区水体均偏负30%以上;吴起鑫等[31]对长江三峡水库坝前水体的研究表明,和天然河流相似,DIC浓度在夏季小于冬季,而δ13CDIC值则是丰水期较枯水期偏负;赖冬荣等[32]对南昌市湖泊水体的研究表明DIC 浓度在春季高于秋季,δ13CDIC则是秋季更为偏正;Wachniew[33]对波罗的海第二大支流Vistula河的研究发现DIC浓度在高流量期最低,而δ13CDIC值在洪水期为-14.2‰,平水期为-9.6‰;Cai等[34]对密西西比河下游的研究发现DIC浓度在丰水期偏低,而枯水期较高,δ13CDIC则在枯水期偏正. 针对梯级水库开发河流DIC及δ13CDIC的变化,李晓东等[35]对嘉陵江干流梯级水库河段的DIC进行研究发现,在丰水期和枯水期DIC的平均浓度分别为2.02和3.15 mmol/L,δ13CDIC值分别为-8.6‰和-6.0‰;类似的还有刘丛强[36]对沅江水系舞阳河、清水江,贾国东[37]等对珠江流域西江和北江河和梁翠翠[27]对九龙江西溪河北溪的研究,这些研究均表明河流溶解性无机碳同位素的组成存在季节变化特征,DIC浓度在枯水期高于丰水期,冬季的δ13CDIC值偏正,本研究结果与其一致. 然而,前述研究结果与乌江梯级水库[16](洪家渡、东风和乌江渡)和猫跳河梯级[38]研究中δ13CDIC值的季节变化规律不同,乌江梯级和猫跳河梯级的研究结果表明,丰水期水体中δ13CDIC值普遍比枯水期偏正(表3),这可能与梯级水库库龄、河流水化学特征及水库的营养状况有较大关系,澜沧江梯级水库中碳的循环转化机制及累积特征还有待深入研究.
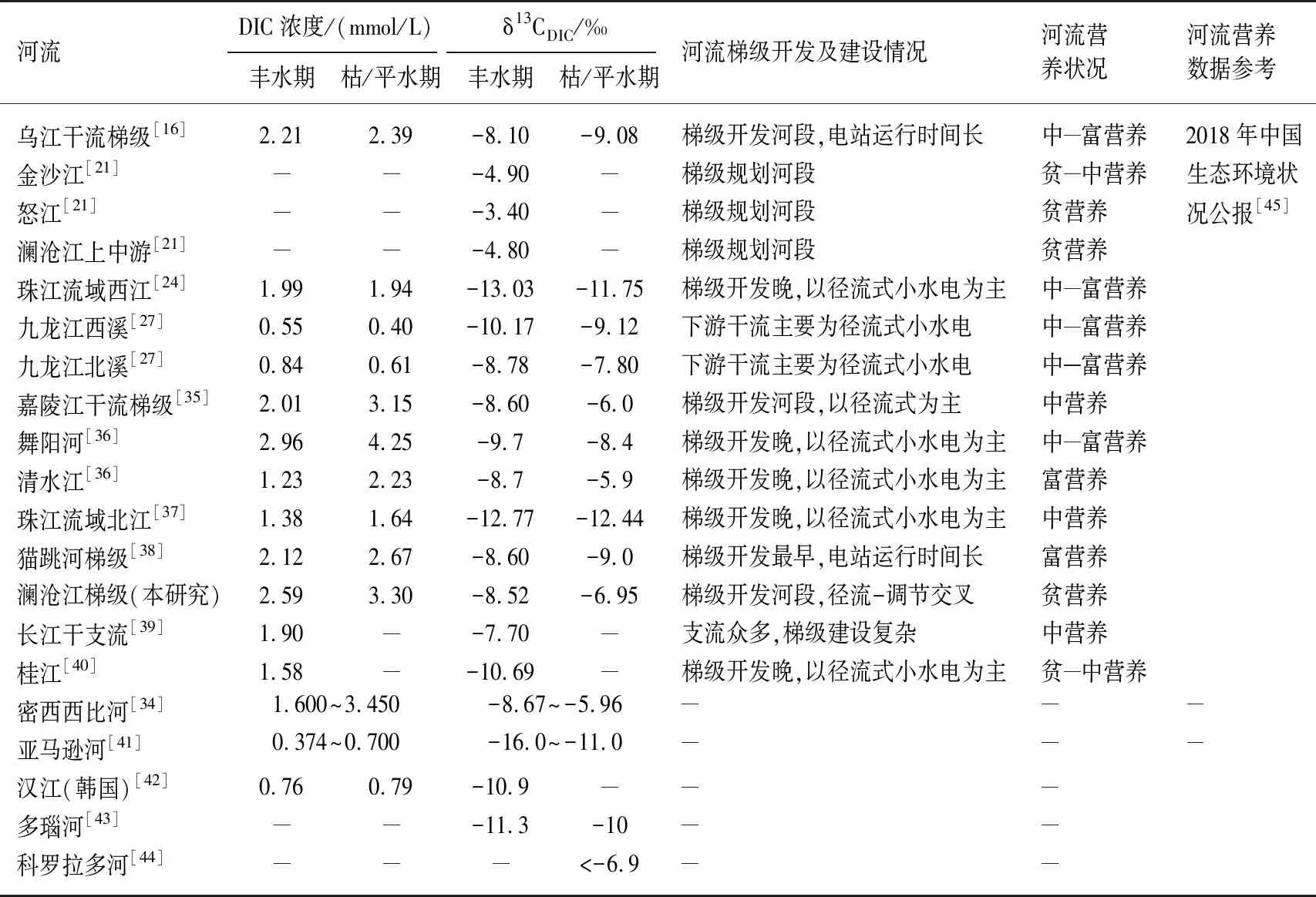
表3 不同流域河流中DIC浓度和δ13CDIC的变化情况
-表示无相关数据.
3.3 澜沧江梯级水库DIC及δ13CDIC变化的驱动因素
3.3.1 流域地质环境因素对梯级河段DIC浓度及δ13CDIC组成的影响 澜沧江云南段河-库系统表层水体中DIC浓度及δ13CDIC呈负相关关系,其中以丰水期表现较为明显(图4a). 这主要可能是一方面在湿季受区域降雨增多影响,澜沧江径流入库水量增大,水体的稀释作用使得DIC浓度明显较低;另一方面,澜沧江云南段干热河谷中夏季温度较高,土壤有机质的分解速率增大,随降雨径流携带进入水体的有机质也会增多,水库形成的缓流开阔水体,有机质作为碳源可以促进表层水体中浮游植物的光合作用,在将无机碳转化为有机碳过程中富含12C的无机碳会优先被藻类吸收,一般会导致表层水体中δ13CDIC值偏正[27]. 然而,澜沧江纵向岭谷区山高坡陡,梯级上中游河段土壤侵蚀严重,样品采集期间正值雨季,大量陆源有机质汇入河道,在其运移过程中氧化分解释放轻碳,湿热环境导致土壤微生物呼吸作用增强、水岩作用充分,硅酸盐矿物溶解速率增大[21, 23]. 与此同时,澜沧江水体在雨季长期处于高浊状态,抑制了浮游植物光合作用中的碳转化效率,这些过程均会使表层水中δ13CDIC向负向偏移. 随着旱季澜沧江流域降雨量的减少和温度降低,土壤有机质分解速率减缓,小湾和糯扎渡等高坝大库的水温稳定分层现象也逐渐消失,水库底部呼吸作用形成的DIC可能受水库水体翻转及循环作用扩散到表层,使得枯水期表层水体DIC浓度增高,进而促使δ13CDIC偏正[38],针对水库垂向δ13CDIC的变化还有待深入研究.
河段中δ13CDIC与DIC的负相关也表明水生呼吸作用对河流的DIC动力学有显著影响,这在以前温带地区的其他研究中也曾报道过[46]. 与此类似,Dubois等[47]的研究表明,与根据1:1混合土壤和碳酸盐来源的DIC估算的理论δ13CDIC值相比,密西西比河流域下游的δ13CDIC(-11.5‰~-7.0‰)明显偏负,并将其归因于有机质的呼吸作用,在密西西比河下游的无机碳库中加入了13C的耗尽碳. 土壤呼吸的CO2作为贫13C来源之一也具有一定的影响,特别是在8月份强降雨时期,强降水可能冲走澜沧江汇水区与让孔隙中积聚的高13C 消耗CO2,并导致河道内13C整体下降. 这种效应在澜沧江中下游的亚热带/热带地区可能更明显,因为这些地区的土壤CO2水平通常高于温带与高山地区[47]. 河-库连续体中水化学参数对DIC浓度和δ13CDIC也有显著影响,pH与水体δ13CDIC的关系研究表明(图4b),碳酸盐岩和硅酸盐岩化学风化对δ13CDIC也有一定影响. 同时,澜沧江河湖连续体中的DIC动力学也可能受到碳酸盐风化作用的影响[48],碳酸盐的风化过程会释放Ca2+和/或Mg2+. 研究发现,δ13CDIC与研究区段Ca2+和Mg2+浓度之间存在一定的相关关系,δ13CDIC值的波动可能与Ca2+和Mg2+浓度变化的过程相同. 一般而言,风化作用作为是水域无机碳通量的主要驱动因素,尤其在低流量条件下具有显著影响[48-49]. 在本研究中,枯水期期间所有水库的δ13CDIC和Ca2+/Mg2+浓度偏高可能是高δ13CDIC来源(如土壤碳酸盐风化)的DIC比例增加所致.
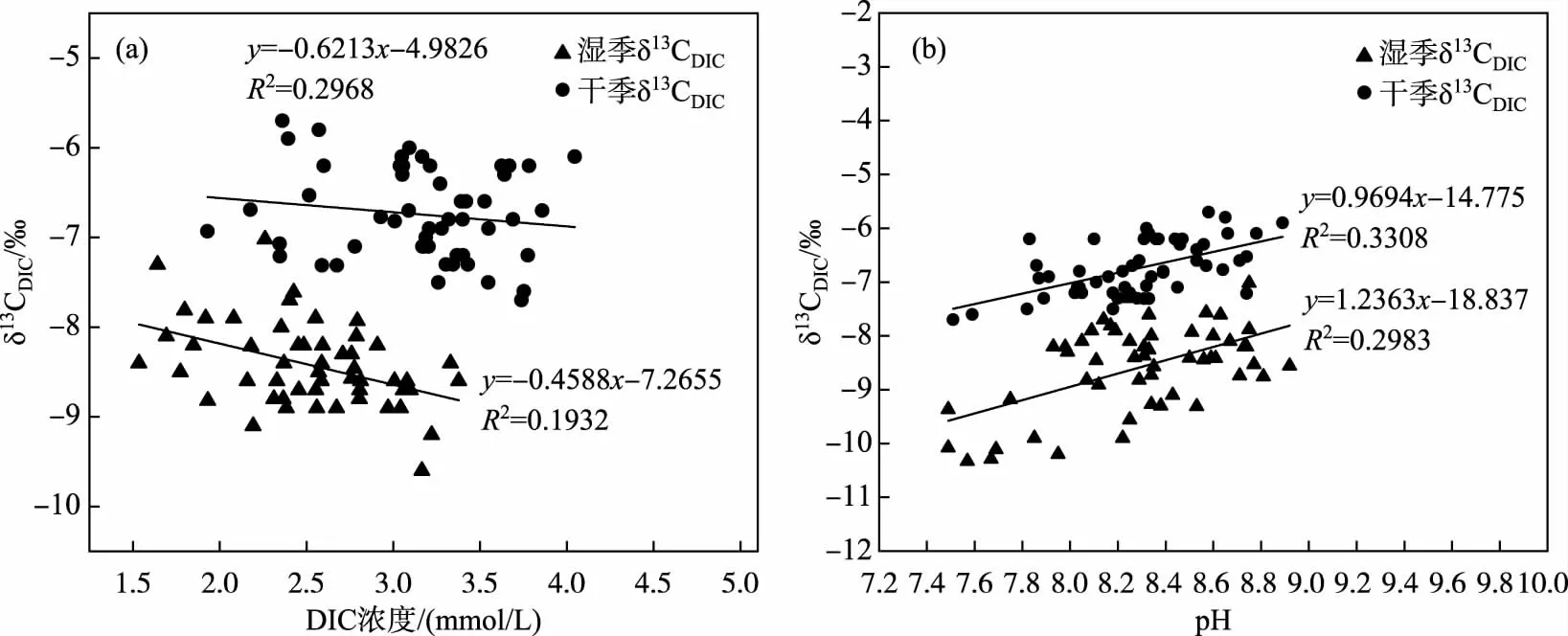
图4 研究区表层水体不同季节DIC浓度和pH与δ13CDIC值的相关关系Fig.4 The correlation between DIC concentrations, pH and δ13CDIC in surface waters of the study area in different seasons
3.3.2 生物地球化学机制驱动下河-库连续体中DIC浓度与δ13CDIC组成的变化 水库δ13CDIC的主要生物地球化学因素包括水库生产力与有机质的分解、水库水体与大气CO2的交换程度、入库水体的DIC碳同位素组成、DIC在水库中的滞留时间等[16]. 研究表明,源于水库所在流域土壤有机质分解形成的DIC具有最负的δ13CDIC值,而流域碳酸盐风化形成的DIC的δ13CDIC值最偏正. 对水-气界面的CO2交换而言,大气CO2溶于水中形成的DIC,其δ13CDIC值约为1.0‰~2.0‰,对水体δ13CDIC值改变较小. 对淡水湖泊碳循环过程的研究[50]表明,表层水体中浮游植物的光合作用和呼吸作用对DIC浓度和δ13CDIC变化相互抵消,所以暂不认为其为控制δ13CDIC变化的主要因素. 水体入库水体DIC的碳同位素组成主要源于河流径流携带和地下水交换过程,而库区周边土壤有机质的分解释放的CO2和流域岩石风化也是DIC的一个主要来源.
DIC滞留时间是水库DIC总量与损失速度的比值,其大小对水体δ13CDIC值有重要影响[51]. 梯级水库中DIC总量主要取决于DIC浓度和水库库容,而DIC损失量包括水库水体与大气CO2交换速度、碳酸盐矿物的沉淀和水库生物量高低等. 对于澜沧江梯级水库来说,水库DIC浓度与上游河段河水较为相近,δ13CDIC值差别也不大. 由于梯级水库的联合运行调度复杂,当前研究还无法得出澜沧江梯级水库群DIC准确的滞留时间,但通过水库河段与上游河段DIC浓度的对比可以看出,调节性能大的水库DIC滞留时间与水库水体δ13CDIC有较好的比例关系. 水库水体与上游河段DIC浓度差别越大,水库中δ13CDIC值变化也越明显,如小湾和糯扎渡水库.
受澜沧江梯级水库联合调度及电站错峰调节的影响,冬季河流处于蓄水期,水库水面面积增加,库中水体的停留时间变长,有利于水体中DIC与大气中CO2之间建立同位素平衡,12C-CO2优先释放进入大气,从而使水库水体中δ13CDIC值向正向偏移. 同时,由于枯水期降水量减少,水流变缓,水体透明度提高,滇西地区适宜的温度适合浮游植物生长,光合作用大于呼吸作用,DIC中12C优先被合成为有机质,同样会使水体中δ13CDIC值向正向偏移,这也是澜沧江干季δ13CDIC值高于湿季的原因.
3.3.3 梯级水电开发对河流DIC浓度及δ13CDIC组成的影响 国内外环境水利学者对大型河流上梯级水利工程建设对径流、水温及常规水质的累积影响已经有诸多报道,但包括水库自身调蓄能力、库龄、运行调度方式、水体停留时间、地质环境条件、库区周边的土地利用及植被覆盖情况对梯级河流物质循环的影响效应随着河流径流变化适应性利用和水库温室气体的研究,最近几年才开始得到涉水工程学界的重视[16-17,27]. 澜沧江云南段梯级水库的建设,改变了河流的水动力条件,已建成的7个水库形成了首尾相接的水库群,导致天然河段基本消失,河流流速减缓,生源物质及泥沙等被大坝截留在库中逐渐沉积或生物转化. 猫跳河流域梯级的研究表明,水库的营养水平越高,生物成因的DIC浓度贡献就越大,从而水体内的δ13CDIC值越偏正[38].
澜沧江云南段河-库连续体中DIC浓度在河流纵向上呈现复杂变化趋势,与乌江流域的研究结果存在较大差异,这可能主要是澜沧江梯级水库在河流纵向空间上建设时间的无序性所导致的[16,35,38]. 云南段现已建成的自上而下“哑铃型”梯级水库布设模式也可能意味着其在碳循环累积影响并不严重,上游多年调节型高坝水库(小湾)及下游多年调节型高坝水库(糯扎渡)中相对较高的DIC浓度和偏正的δ13CDIC值随水库下泄水体分别进入其下一级水库(漫湾和景洪)的过程中,并不一定会在空间上对其产生较强的累积影响,相反DIC浓度可能会随着水流的稀释作用呈现沿程坦化或者衰减,因此导致高坝大库下游的梯级中DIC的浓度并没有上游高,δ13CDIC值也不会明显偏负. 同时,本研究中发现,水库水温与DIC浓度呈负相关,即位于下游河段热带和亚热带的水库(糯扎渡和景洪),河流水温越高其DIC浓度越低. 以水库为研究单元,发现水体中DIC浓度为小湾 > 苗尾 > 功果桥 > 漫湾 > 大朝山 > 景洪 > 糯扎渡,而δ13CDIC值则几乎相反,为糯扎渡 > 景洪 > 大朝山 > 漫湾 > 苗尾 > 功果桥 > 小湾,这表明澜沧江梯级水库δ13CDIC值的变化可能是水库水面面积及扩散系数、电站调节方式和调度运行共同作用的结果.
从时间分布来看,由于澜沧江云南段所建梯级水库水体中DIC浓度和δ13CDIC值的季节变化特征与许多自然河流和三峡水库坝前水体表现出一致性[35],而与乌江和猫跳河等较早建设梯级的结果差别明显[16,38]. 除地质背景差异外,这可能与澜沧江梯级水库群建库时间短、联合运行条件下调度方式复杂和水文条件多变密切相关,梯级电站的“水库效应”还没体现出来. 已有的研究表明[51-52],水库水体停留时间越长,水体中营养物质生物地球化学循环的转化过程越强烈,其演化和湖沼化水平也越高;澜沧江云南段的梯级水库因停留时间较短,水库内转化过程相对较弱.
4 结论
本研究对澜沧江云南段河-库连续体丰水期和枯水期水体中溶解性有机碳及其同位素的时空变化特征、物源特征及影响因素进行了分析. 研究结果表明,澜沧江水体DIC浓度及δ13CDIC值存在显著的季节变化特征,均为冬季(枯水期)高于夏季(丰水期). 丰水期DIC浓度明显低于枯水期,主要可能是夏季上游流域降雨增多、入库水量增大所产生的水体稀释作用导致的;丰水期δ13CDIC值偏负的原因可能是夏季澜沧江纵向岭谷区湿热环境导致土壤微生物呼吸作用增强、水岩作用充分、碳酸盐矿物溶解速率增强;丰水期澜沧江水体处于高浊状态,抑制了浮游植物光合作用过程中碳的转化效率. 梯级水库的拦截后会导致河流水化学性质发生改变,但本研究结果与前人研究所证实的自然河流的季节变化特征相似,而与天然湖泊、库龄较长的水库不同,这可能与澜沧江梯级水库纵向空间上建设时间无序、水库累积影响不显著、联合调度运行复杂和水文条件改变等因素有关,综合表明澜沧江梯级水库群水体的“水库效应”并不明显.
