谷子SiWRKY03基因的分子特征与表达分析
2020-01-08宋振君李志勇王永芳董志平
宋振君,李志勇,王永芳,刘 磊,白 辉,董志平
(1.河北省农林科学院 谷子研究所,国家谷子改良中心,河北省杂粮重点实验室,河北 石家庄 050035; 2.河北师范大学 生命科学学院,河北 石家庄 050024)
WRKY是植物中最大的一类转录因子家族之一,1994年在甘薯中发现第一个WRKY转录因子SPF1,随后在拟南芥[1]、水稻[2]、大麦[3]、黄瓜[4]、玉米[5]、棉花[6]、番茄[7]和小麦[8]中逐步有很多相关研究。WRKY家族成员都含有由60个氨基酸组成的WRKY结构域,其N端有保守的WRKYGQK序列(WRKY因此得名),C端包含锌指结构C2H2或C2HC[9]。
WRKY家族成员广泛地参与植物的生长发育与形态建成[10]。如AtWRKY44(即TTG2)是拟南芥中首次发现的与表皮毛和种皮发育相关的WRKY转录因子[11];AtWRKY6在拟南芥衰老的叶片内表达量显著提高,表明其参与叶片的衰老过程[12]。水稻中OsWRKY23的表达主要在根和衰老叶片中,说明其参与根和衰老叶片的表达调控[13];OsWRKY13、OsWRKY23、OsWRKY71、OsWRKY12、OsWRKY43、OsWRKY55与OsWRKY86 7个蛋白质在苗期表达量较低,随叶片的生长表达逐步增加,推测它们在叶片的正常生长过程中发挥作用[14]。
除了在植物正常生长中发挥作用外,WRKY转录因子还参与了植物的抗病过程。OsWRKY22敲除的水稻株系对稻瘟病菌感病,而过表达可增强稻瘟病抗性[15];OsWRKY71提升了转基因水稻对白叶枯病的抗性[16];过表达PtrWRKY89增加了杨树对黑斑病的抗性[17];TaWRKY70沉默增强了小麦对条锈菌的感病性[18]。WRKY转录因子通过其结构域与靶基因启动子区域的(T)(T)TGAC(C/T)序列(W-box)特异性结合,以调控下游基因的转录[19]。如OsWRKY53超表达的转基因水稻中,OsWRKY53特异性地结合在W盒序列元件上,诱导PBZ1、PR5、PR14、几丁质酶与过氧化物酶等防御相关基因的表达,提高水稻对稻瘟病的抗性[20];较未转基因水稻相比,OsWRKY80超表达水稻增强了病程相关基因ZB8、PBZ1的表达,表现出对稻瘟病较好的抗性[21];ZmWRKY79过表达的玉米原生质体依赖W-box或WLE顺式元件,增加了萜类植保素生物合成相关基因的表达,提高了植株抗生物胁迫能力[22]。上述结果说明,WRKY转录因子可以诱导防御相关基因的高水平表达,从而正调控水稻对稻瘟病的抗性反应。然而,谷子中关于WRKY转录因子在抗病反应中的相关研究尚无报道。
谷锈病(Uromycessetariae-italicaeYoshino)是谷子上的重要病害,常年减产10%~30%,流行年份植株倒伏、颗粒无收,严重影响着谷子的稳产和高产[23]。生产中利用抗锈品种是最经济有效的防治方法。但抗锈品种应用中易出现抗性丧失现象,因此,克隆抗锈基因、揭示抗锈机理对快速培育抗锈品种,延长品种使用寿命具有重要意义。WRKY是一类对植物生长发育、抗病反应具有重要作用的转录因子,拟南芥中AtWRKY29/22与水稻中同源基因OsWRKY03都参与了植物的防御反应[24-25],笔者对其在谷子中的同源基因SiWRKY03进行了系统研究,包括利用生物信息学软件分析了其生物学特征,采用Real-time PCR技术检测了SiWRKY03转录因子在谷子不同组织部位和抗谷锈病过程中的表达丰度变化,为探索WRKY成员的功能,了解谷子抗病机理提供试验数据。
1 材料和方法
1.1 试验材料及试剂
试验植物材料为谷子抗病材料十里香和感病材料豫谷1号,供试菌株为谷锈菌强毒性小种A57的单孢菌系93-5,均由河北省农林科学院谷子研究所植保室保存。十里香对谷锈菌单孢菌系93-5的侵染表现抗病反应,豫谷1号对93-5的侵染表现感病反应。
TRIzol试剂、cDNA第一链合成试剂盒(RevertAidTMFirst Strand cDNA Synthesis Kit)购自赛默飞世尔(中国)科技公司;Real-time PCR使用的TB GreenTMPremix EX TaqTMⅡ购自TaKaRa;引物合成由苏州金唯智生物科技有限公司完成。
1.2 试验方法
1.2.1 试验材料处理 谷子发育过程中的不同组织部位取材:选取谷子品种豫谷1号孕穗期的根、茎、叶、穗4个部位进行样品采集,锡箔纸包裹置于液氮速冻后,于-80 ℃保存。叶片的谷锈菌接种处理与取材:选取21 d(四-五叶期)长势一致的十里香和豫谷1号的幼苗,采用新鲜的谷锈菌单孢菌系93-5制备的孢子悬浮液(浓度为1.0×105个/mL)对幼苗进行喷雾接种,分别于接种后0,12,24,36,48,72,96,120 h 8个时间点剪取叶片,锡箔纸包裹置于液氮速冻后,于-80 ℃保存。
1.2.2 谷子SiWRKY03蛋白生物信息学分析 利用Phytozome植物基因组网站(https://phytozome.jgi.doe.gov/pz/portal.html)下载的谷子豫谷1号参考基因组(Setariaitalicav2.2),查找SiWRKY03(基因ID:Seita.5G233100)的CDS序列与蛋白质序列。利用NCBI的Conserved Domain Database(https://www.ncbi.nlm.nih.gov/cdd)预测SiWRKY03蛋白质的结构域;利用ExPASy-ProtParam tool(https://web.expasy.org/protparam/)对SiWRKY03蛋白质的理化性质进行分析;利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL(https://swissmodel.expasy.org/)软件预测SiWRKY03蛋白的二级、三级结构;将SiWRKY03蛋白质序列在NCBI 中进行Protein Blast比对(https://blast.ncbi.nlm.nih.gov/Blast.cgi),按照相似性程度高低选择来自10个物种的WRKY03的同源基因的蛋白质序列,采用Clustal X 2.0进行多重序列比对,利用MEGA 6以邻近法(Bootstrap设为1 000)构建系统进化树,分析其亲缘关系的远近。
1.2.3 谷子中SiWRKY03的实时荧光定量PCR检测与分析 采用TRIzol试剂提取不同处理条件下取材的谷子样品的总RNA,按照cDNA第一链合成试剂盒的说明书反转录得到cDNA,作为Real-time PCR的模板。
根据SiWRKY03的CDS序列,采用Primer Premier 5.0设计Real-time PCR引物:SiWRKY03-F(5′-AGAGCCAGCTGAAGAAGGTG-3′)/SiWRKY03-R(5′-CTTCATGCTGCTGCACTTGT-3′);选用谷子肌动蛋白基因Actin(Seita.8G043100.1)作为内参,设计Real-time PCR引物:Actin-F(5′-CGCATATGTGGCT CTTGACT-3′)/Actin-R(5′-GGGCACCTAAATCTCTCT GC-3′)。Real-time PCR反应体系为20 μL :10 μL TB GreenTMPremix Ex TaqTMⅡ、0.8 μL上下游引物、2 μL cDNA模板和6.4 μL灭菌双蒸水。反应程序为:95 ℃预变性30 s;95 ℃变性15 s,60 ℃退火30 s,72 ℃延伸32 s,共40个循环收集荧光信号。每个反应重复3次,Ct值取平均值,采用2-ΔΔCt法计算基因在不同样品中的相对表达量(表达倍数的阈值设定为>2,<0.5)。
2 结果与分析
2.1 SiWRKY03蛋白的结构特征
SiWRKY03基因的CDS序列全长为1 137 bp,共编码378个氨基酸(图1)。195-251氨基酸序列(WRKYGQK)为WRKY转录因子的保守结构序列,是一个DNA结合结构域,能特异性的与目标基因启动子区域的(T)(T)TGAC(C/T)序列(Wob)结合,以调控基因表达,主要参与植物的抗病防御、衰老和根毛的发育。

下划线代表SiWRKY03蛋白的保守结构域。 The underline represents the conservative domain of SiWRKY03 protein.
2.2 SiWRKY03基因的生物信息学分析
2.2.1SiWRKY03基因编码蛋白质的理化性质 利用在线ExPASy-ProtParam tool对SiWRKY03编码蛋白质的理化性质进行分析,预测蛋白质分子质量为39.73 ku,pI为5.91,平均疏水性系数为-0.498,属于亲水性蛋白质。SiWRKY03基因编码378个氨基酸,其中丙氨酸(Ala)含量最高(17.7%),天冬酰胺(Asn)、异亮氨酸(Ile)和色氨酸(Trp)含量最低(分别是1.1%),带负电荷氨基酸(天冬氨酸(Asp)+谷氨酸(Glu))有52个,带正电荷氨基酸(精氨酸(Arg)+赖氨酸(Lys))46个。
2.2.2 SiWRKY03编码蛋白的二级、三级结构预测 利用SOPMA软件对SiWRKY03编码的蛋白质进行二级结构预测(图2-A),发现该蛋白的二级结构中无规则卷曲(Random coil)占52.65%、α-螺旋(Alpha helix)占34.13%、延伸链(Extended strand)占6.88%、β-转角(Beta turn)占6.35%。由此可见,SiWRKY03蛋白质的最大结构元件是无规则卷曲,最小的元件为β-转角。运用SWISS-MODEL软件对SiWRKY03蛋白质的三级结构进行预测与显示(图2-B),并将结果与二级结构预测结果进行比对,结果较为一致。
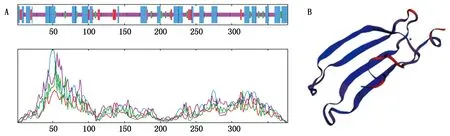
A.二级结构;B.三级结构。 A. Secondary structure; B.Tertiary structure.
2.2.3 SiWRKY03同源性分析 把SiWRKY03编码的氨基酸序列在NCBI上进行Protein Blast比对,按照序列相似性程度选择了来自玉米、水稻、小麦、青稞、高粱、糜子、哈氏黍、二穗短柄草、Dichantheliumoligosanthes和大豆中的WRKY03同源基因的蛋白质序列构建系统进化树(图3)。结果表明,SiWRKY03与单子叶植物糜子(Panicummiliaceum,RLM93064.1)和哈氏黍(Panicumhallii,PAN29607.1)氨基酸序列的同源性最高(99%),其次是Dichantheliumoligosanthes(OEL17299.1),同源性为96%;与双子叶植物大豆(Glycinemax,XP_003548638.1)同源性最低,仅为28%。
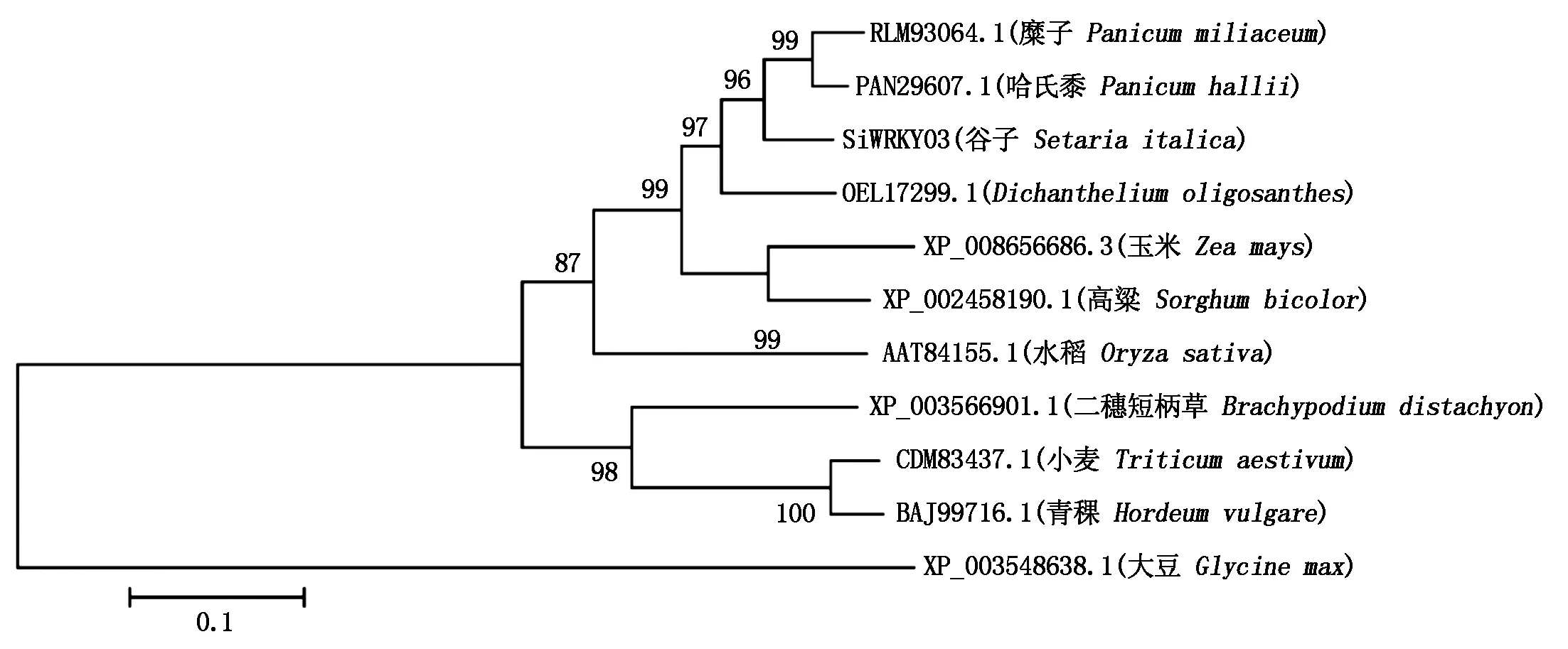
图3 SiWRKY03的系统进化树分析Fig.3 The phylogenetic tree analysis of SiWRKY03
2.3 SiWRKY03基因的组织表达分析
为了解SiWRKY03基因在谷子不同组织部位中的表达情况,采用Real-time PCR技术检测了该基因在谷子豫谷1号孕穗期的根、茎、叶和穗4个部位的表达量。结果如图4所示,SiWRKY03在谷子孕穗期的根、茎、叶和穗中均有表达,但主要在根中表达,在其他组织部位中表达水平较低,穗部最低。其中,SiWRKY03在根中的表达量是茎的4.34倍,是叶的3.57倍,是穗的12.50倍。豫谷1号孕穗期根、茎、叶和穗部的转录组数据显示,SiWRKY03在根中的表达量最高,是其他3个部位表达量的5.81,3.80,5.99倍,与Real-time PCR结果基本一致。
2.4 生物胁迫下SiWRKY03基因的表达分析
采用Real-time PCR检测分析了SiWRKY03基因在谷子抗病材料十里香和感病材料豫谷1号接种谷锈菌后0,12,24,36,48,72,96,120 h的表达情况。结果如图5所示,SiWRKY03在抗病和感病2种反应中的表达模式不同。抗病反应中,SiWRKY03的表达量在12 h上调并达到最大表达量,是起始表达量的3.47倍。在36 h同样上调表达,表达量为起始表达量的2.16倍,其他时间点SiWRKY03的表达量无显著变化。在感病反应中,SiWRKY03的表达量在所有的时间点均未发生显著的变化。推测SiMYB003与抗病相关,且在抗病反应的早期起正调控作用。

图4 SiWRKY03在谷子的不同组织部位的表达分析Fig.4 Analysis of expression of SiWRKY03 gene at different tissue of foxtail millet
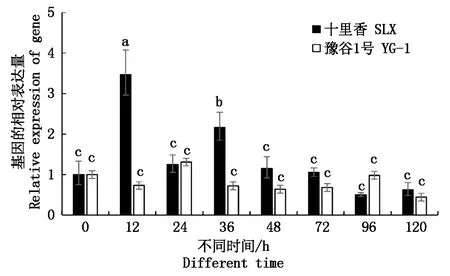
不同小写字母表示差异显著,P<0.05。 Different lowercase indicate significant different at 0.05 level.
3 结论与讨论
本研究利用Real-time PCR分析SiWRKY03在豫谷1号中不同组织部位的表达情况,结果显示,在根、茎、叶和穗中均有表达,在根中表达量最高,推测其参与植株根部的发育过程。邢国芳等[26]采用生物信息学方法,对谷子基因组中WRKY家族进行鉴定与分析,得到103个WRKY转录因子,并对其在不同组织中的表达进行检测,结果发现,其中80个SiWRKY基因在根中的表达量很高;SiWRKY51和SiWRKY31在叶片中具有高表达量;SiWRKY86和SiWRKY87在茎中高表达;SiWRKY4、SiWRKY6、SiWRKY34、SiWRKY54、SiWRKY57和SiWRKY67在穗中特异性表达,说明WRKY家族基因具有组织表达特性。其中,SiWRKY03基因在根中特异性表达[26],与本研究结果一致,猜测SiWRKY03可能在根的生长发育中起到一定的作用。本试验结果对了解WRKY 转录因子的功能进一步积累了重要数据。
同样,本研究还分析了SiWRKY03在响应谷子抗谷锈菌胁迫反应中的表达情况。SiWRKY03在谷子十里香抗锈病反应中上调表达,在豫谷1号感锈病反应中表达量无显著变化,2种表达模式的不同提示基因可能与抗病相关,且为正调控因子。在其他物种中已有此基因的同源基因的相关研究,在水稻中的OsWRKY03超表达会导致OsNPR1以及其他几种病程相关基因(如OsPR1b、苯丙氨酸-氨解酶ZB8和过氧化物酶POX22.3)的表达量的提升,表明OsWRKY03对于水稻抗真菌是正调控因子[25];OsWRKY03在拟南芥中的同源基因AtWRKY29/22编码的蛋白,可以被MAP激酶信号级联途径激活,过表达可使植株对细菌和真菌产生抗性[24]。这2个研究结果与本试验结果一致,由此推断SiWRKY03基因在谷子抗锈病反应过程中起正调控作用,但抗病是一个复杂的过程,具体的作用机制有待进一步的研究。
