小麦雄性不育系BS366和FA-101的配合力及杂种优势分析
2020-01-05苑少华李艳梅白建芳段文静谭照国刘子涵赵昌平张风廷陈兆波张立平
苑少华,李艳梅,白建芳,段文静,谭照国,3,刘子涵,王 娜,赵昌平,张风廷,陈兆波,张立平
(1. 北京市农林科学院杂交小麦工程技术研究中心,北京 100097;2. 杂交小麦分子遗传北京市重点实验室,北京 100097;3. 长江大学生命科学学院,湖北荆州 434025)
小麦是我国重要的粮食作物之一。随着社会经济的发展以及人口的快速增长,小麦的需求量也呈增加趋势。在耕地面积不断减少的形势下,小麦总产量的提升只能通过单产的大幅度提高来实现[1]。杂种优势利用是提高作物产量的有效途径之一[2],而小麦还未大规模实现杂种优势利用[3]。近年来,我国基于小麦光温敏不育系创建了“二系法杂交小麦应用技术体系”,由于其操作简单、恢复源较广、制种成本低等优势,该杂优利用体系成为杂交小麦应用的主要途径[4]。在生产中应用的BS型小麦光温敏不育系为“京麦”系列杂交小麦品种的母本,其新一代不育系的光敏特性显著,大幅提高了不育系的稳定性。F型三系不育系材料是我国新发现的不育源,是一种新型普通小麦细胞质雄性不育系[5],而且其育性具有一定的光温敏感特性[6]。
小麦杂种优势大面积利用中存在诸多制约因素,其中强优势组合的选配是关键[7]。强优势组合的选配主要是通过亲本配合力分析对杂交种进行亲本选择和组合评价[8]。小麦产量构成的有效穗数、穗粒数和千粒重三要素中,千粒重的遗传力最大[9-10]。通过分析不育系及恢复系主要农艺性状的配合力,对筛选强优势杂交组合、提高育种效率具有非常重要的意义。前人对于杂交小麦杂种优势利用及配合力分析也有报道,但尚无针对两系或F型雄性不育系的配合力及杂种优势比较的相关研究。前期李晓燕[11]对光温敏二系小麦BS210和BS366的杂种优势和配合力进行了评价,陈 林[12]、温辉芹[13]和任永康[14]等仅对F型不育系杂种优势进行了评价,但并未对配合力进行深入分析。本研究利用光温敏雄性不育系BS366和F型雄性不育系FA-101及其杂交组合对其产量性状的杂种优势和配合力进行分析,探索强优势组合的选配规律,以期对二系及三系杂交小麦强优势组合的组配提供理论 支持。
1 材料和方法
1.1 试验材料
供试材料为两系光温敏核雄性不育系BS366、F型三系雄性不育系FA-101及16个恢复系材料(川麦56、济麦12、绵阳35、川育21、凤麦24、07品151-180、川麦43、川农16、蜀麦482、川麦52、Syn53、太原608、PI601477、PI17901、PI486146和PI598210,分别用P1~P16表示)。
1.2 试验方法
2018年4月以BS366和FA-101为母本,与16个父本材料进行杂交,2018年9月下旬将杂交收获的F0代材料种植于北京(N 39°54′, E 116°18′)生态区,随机区组设计,三次重复,两行区, 1.5 m行长,行距25 cm,株距5 cm,人工点播,田间管理同大田生产。2019年5月对亲本、F1代株高、穗长、单株穗数、小穗数、穗粒数、千粒重和单株粒重进行调查。
单株粒重=单株穗数×穗粒数×千粒重/ 1 000
1.3 数据分析和处理
配合力分析采用不完全双列法杂交[15],各供试亲本的一般配合力(GCA)按gi=xi-X计算;各参试组合的特殊配合力(SCA)按sij=xij-X-gi-gj计算。其中xi和xj分别表示i亲本和j亲本所配组合的性状平均值,X为所有组合的总平均值。对调查的亲本7个性状值以及7个性状的GCA和SCA在DPS软件下进行分析。
超中亲优势=(F1-PA)/PA×100%,PA为双亲平均值;超亲优势=(F1-P)/P×100%,P为较优亲本值;对照优势=(F1-CK)/CK×100%,CK为对照值。
2 结果与分析
2.1 亲本及杂交组合的配合力分析
2.1.1 杂交组合配合力方差分析
对具有的7个相同父本(P1、P3、P5、P9、P12、P14和P15)的14个杂交组合的7个性状进行方差分析表明,各性状在组合间均存在极显著差异(P<0.01)或显著差异(P<0.05),而区组间除株高和穗长外均不显著;父本间株高的GCA值差异显著,母本间株高和小穂数GCA值差异极显著,其余性状均不显著;组合间各性状SCA值均极显著(表1),因此可进一步分析亲本的GCA和组合的SCA效应。
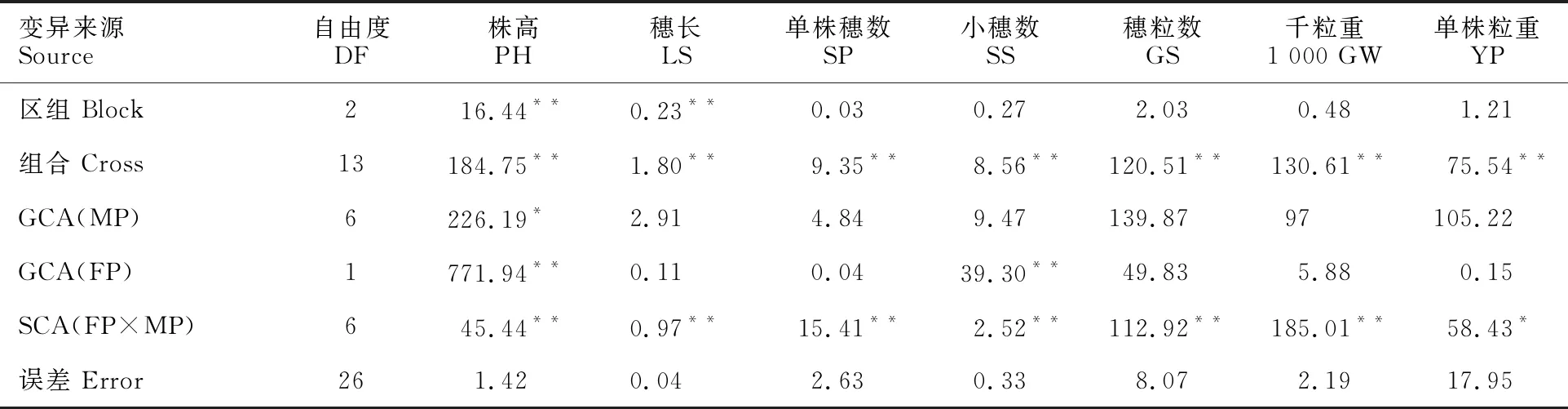
表1 7个性状的方差分析(F值)
2.1.2 亲本的GCA效应分析
GCA指一个亲本与多个亲本杂交后杂种后代在某个性状上的平均表现,主要由基因的加性效应决定,是可以稳定遗传的部分[16]。从表2可以看出,同一亲本的不同性状间和同一性状的不同亲本间GCA值都有较大差异,说明不同亲本在性状上的加性基因效应值大小不同。
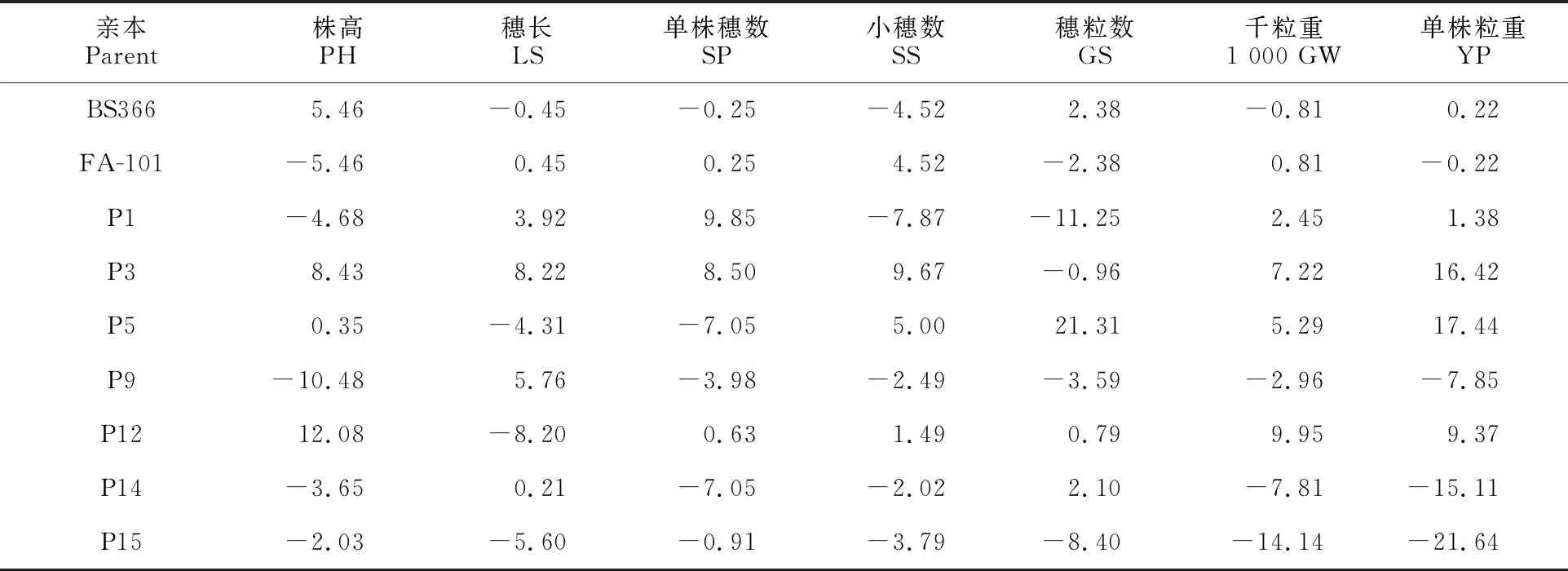
表2 9个小麦亲本7个性状的一般配合力(GCA)效应值
在7个父本中,P9、P1、P14和P15的株高GCA为负值,相对效应较大,说明这4个材料在矮秆育种中有一定的利用价值;穗长GCA值较大的有P3、P9和P1;小穗数GCA值较大的有P3和P5;单株粒重GCA值较大的有P5、P3和P12。此外,在单株粒重GCA值为正值的P1、P3、P5和P12的4个亲本中,在单株穗数、穗粒数、千粒重等3个产量性状中均至少具有2个或2个以上性状存在GCA正向效应值(表3),表明本研究中影响产量的最终因素并不是某单一性状特别突出而造成的,而是产量要素共同作用的结果。
2.1.3 杂交组合产量三要素和单株粒重的SCA效应分析
SCA是由非加性效应产生的,很难在后代中稳定遗传,但亲本间的特殊配合力效应大,有利于选育出该性状表现优异的突破性杂交小麦组合。从表4中可以看出,不同组合单株粒重的SCA值有很大差异,其变化范围为-21.99~21.99。其中SCA值较大的组合有BS/P9、FA/P1、FA/P12和FA/P15,其单株穗数、穗粒数和千粒重中至少有2个或2个以上性状的SCA值表现出正向效应;SCA值较小的组合有FA/P9、BS/P1、BS/P12和BS/P15(表3)。

表3 14个杂交组合7个性状的特殊配合力(SCA)效应值及其多重比较分析
2.2 杂交组合的杂种优势分析
从23个杂交组合F1代的杂种优势(表4)看,7个性状超中亲优势组合比例为56.52%~100%,超中亲优势明显,且7个性状的超中亲优势均值均为正;超高亲优势组合比例为17.39%~65.22%,穗长、单株穗数、千粒重和单株粒重的超高亲优势均值为正;超对照优势组合比例为30.43%~95.65%,穗长、单株穗数和小穗数的超对照优势均值为正。
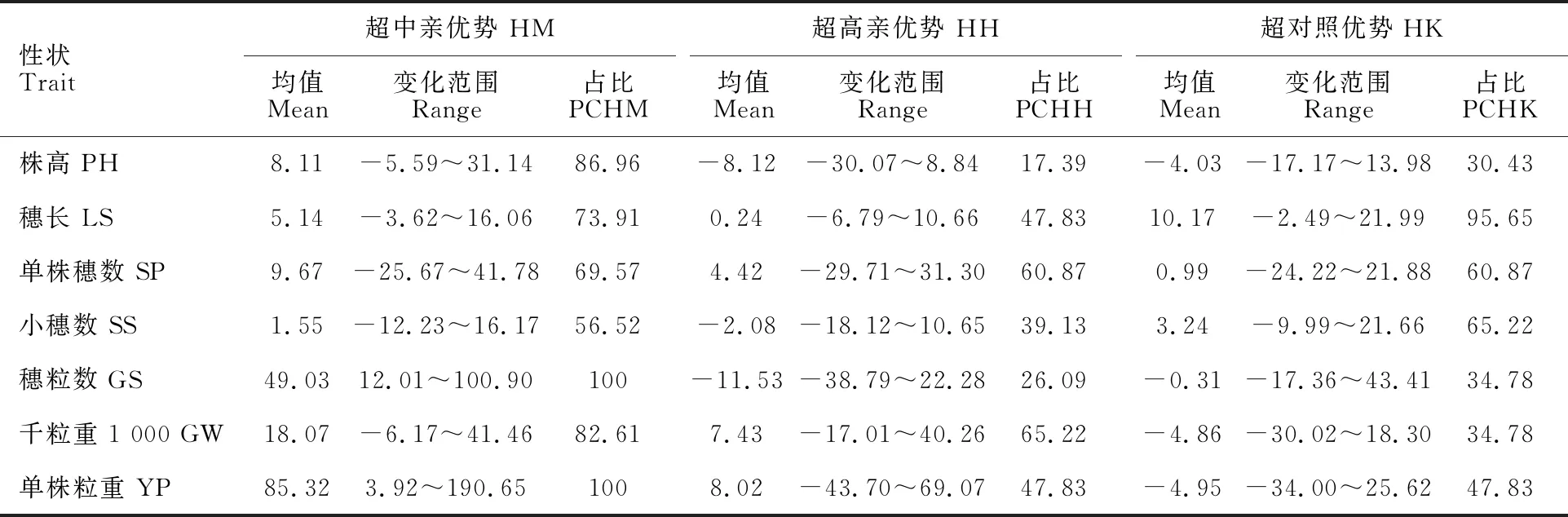
表4 杂交组合F1代各农艺性状的中亲优势和超亲优势
从23个杂交组合中筛选出11个组合(6个BS366组合,5个FA-101组合),其单株粒重超对照优势为正,范围为2.73%~25.62%,株高超对照优势为-13.53%~10.31%(表5)。其中BS/P2、BS/P5、BS/P6和FA/P4等4个组合的单株粒重超对照优势均大于12%,且株高均低于 对照。
此外,除BS/P2、BS/P5、BS/P7和FA/P1以外的7个组合在单株穗数、穗粒数、千粒重等3个产量性状中至少有2个或2个以上的杂种优势为正,表明本研究中影响产量的最终因素并不是某单一性状特别突出而造成的,大多是产量要素共同作用的结果。
另外发现,父本P3和P5组合的单株粒重均超对照(表5),因此这2个亲本可能作为优良父本与其他不育系进行组合选配,进而发掘新的具有杂种优势的组合。
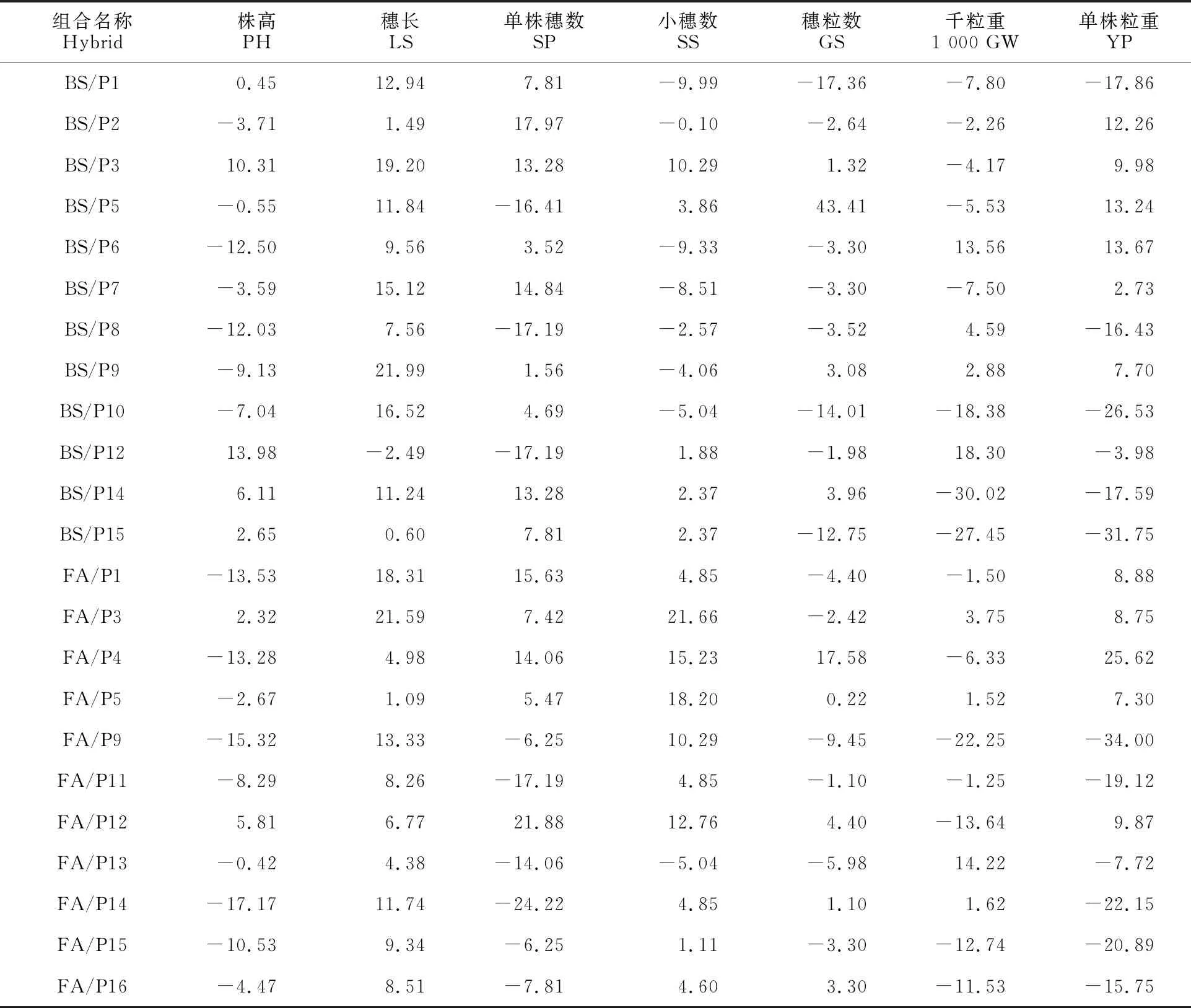
表5 7个农艺性状的超对照优势
2.3 杂种优势与配合力的相关性分析
在小麦杂交育种中,杂种优势与配合力有着密不可分的关系,具有良好配合力的亲本或品系才能组配出更优异的杂交种,因而亲本配合力是选择优良杂交种的重要依据[16]。一个杂交组合的优劣由亲本的GCA和组合的SCA效应共同决定[17]。目前,产量是评价杂种优势的最主要标准之一[18]。
对14个杂交组合的杂种超对照优势值与父本GCA值和组合SCA值的相关性分析表明,杂种优势值与GCA和SCA都有较高的相关性,且与GCA的相关性要大于与SCA的相关性,但父本GCA值与组合SCA值几乎无相关性(表6)。

表6 杂种优势与GCA、SCA的相关性
2.4 BS不育系与FA不育系配合力及后代杂种优势比较分析
2.4.1 BS366和FA-101配合力比较分析
BS366和FA-101在株高、小穗数和穗粒数等3个性状上的GCA差异明显(表2)。BS366单株穗数和千粒重的GCA值低于FA-101,但单株粒重要优于FA-101。因此,在杂交小麦新组合的选育上,应根据不育系的特点,定向选择可以优势互补的恢复系进行组配,有利于获得产量水平较高的强优势组合。
在单株穗数、穗粒数、千粒重和单株粒重等产量相关性状中,相同父本的BS366和FA-101的7对组合SCA值之差的范围分别是6.26~37.92、1.05~38.24、5.15~35.94和1.17~43.99,而且每个性状下均有4对组合的SCA值之差大于10(表3),说明这2种不育系的相同父本组合大多数情况下的特殊配合力有较大差异。
2.4.2 BS366和FA-101杂种优势比较分析
相同父本情况下分别对BS366和FA-101的所有组合的杂种优势进行比较(表5)。结果发现,BS366和FA-101分别与P3、P5、P14和P15的杂交组合的超对照优势比较一致,均为正或负。此外,两个不育系与P1、P9和P12的杂交组合的超对照优势值的正负完全相反,而其余9个父本的组合中均只有1个组合为高恢复性组合(结实率>90%),因此无法进行对比,表明这两个不育系的组合的杂种优势和恢复性存在明显不同。
2.4.3 BS366和FA-101杂种优势相关性分析
对上述相同父本的BS366和FA-101的7个杂交组合间的7个性状的杂种优势进行相关性分析发现,只有株高间的相关性大于0.6,具有较强的相关性;穗长、小穂数、穗粒数和株粒重只具有中度或中度以下的相关性;而单株穗数和千粒重具有负相关性(表7)。此外,分别对BS和FA组合的单株粒重与产量三要素的相关性分析发现,BS366组合中单株粒重与穗粒数和千粒重具有较强的相关性,与单株穗数具有一定的负相关性;FA-101组合中与单株穗数具有极强相关性,与穗粒数和千粒重具有一定的相关性(表8)。因此,BS366和FA-101两种不育系配制的杂交组合,其杂种优势趋势不同,尤其是在单株穗数、穗粒数、千粒重和单株粒重等产量性状方面存在较大差异(表7、表8)。

表7 具有相同父本的BS366和FA-101组合的各农艺性状相关性

表8 BS366组合和FA-101组合的株粒重 与产量三要素的相关性
3 讨 论
3.1 BS及F型不育系优良杂交组合及父本的 筛选
本研究结果表明,影响BS366组合产量的性状顺序为穗粒数、千粒重>单株穗数,影响FA-101组合的顺序为单株穗数>穗粒数、千粒重(表8),两者结果不同,与前人研究结论存在偏差[18-20],其原因应该是试验材料不同导致的差异。同时发现穗粒数的超高亲和超对照优势均为负,且超高亲优势组合仅占26.09%(表4),在所有产量性状里最低,说明不育系育性恢复性偏低影响结实率是二系杂交小麦产量提高的瓶颈。因此,应特别重视对恢复系的选择,进而大幅度提高穗粒数的超亲(或超标)优势,是超高产杂交小麦组合选育的关键,也是难点之一。
本研究通过对杂种优势的分析,筛选出BS/P2、BS/P5、BS/P6和FA/P4等4个高产杂交小麦新组合,其是否能进行生产应用,还需要从双亲花期、株高、制种产量等方面进行试验;父本P3和P5可作为优良核心恢复系使用。
3.2 BS366与FA-101的组合的比较分析
总体上讲,小麦杂种优势利用相对落后,经过几代人努力目前已取得了较大的成绩,但是关于小麦光温敏两系和F型三系不育系利用相同父本配制杂交组合,并比较其杂种优势和配合力的研究鲜有报道。前期苑少华、孙辉等[6,21]对光温敏二系不育系BS366和F型三系不育系的恢复性研究发现,二者都具有不同程度的光温敏特性,但其恢复源不完全一致,其不育及育性恢复机制存在一定差异。本研究发现,BS366和FA-101两个不育系各性状的一般配合力不同(表2),具有相同父本的BS和FA组合间的杂种优势趋势、规律(表5、表7、表8)以及特殊配合力效应值(表3)也大多存在差异,进一步验证了BS和FA属于不同类型的不育系,而且两者提高杂种优势潜力的策略有所不同。
