生物炭在柑橘皮厌氧发酵中作用机理研究
2019-12-06MichaelFagbohungbeBenHerbertLoisHurstHongLiShamsUsmaniKirkSemple
Michael O Fagbohungbe , Ben M J Herbert , Lois Hurst , Hong Li , Shams Q Usmani , Kirk T Semple
(1.Lancaster Environment Centre, Lancaster University, Lancaster LA1 4YQ, United Kingdom; 2. Stopford Energy and Environment, Merseyton Road, Ellemere Port, Chester CH65 3AD, United Kingdom; 3. Ariva Technology, The Heath Business and Technical Park, Runcorn, Cheshire WA7 4EB, United Kingdom)
1 柑橘皮废料处理工艺现状
柑橘皮废料是一种含有纤维素和香精油的木质纤维材料,其中32%~98%的成分是由烷基芳烃合成的柠檬烯。柠檬烯是一种无色液体,有刺激性气味,其沸点是176℃,属于环萜烯。厌氧消化(AD)的研究表明柑橘皮在有机负荷为2~3.5 g-1VS·d-1时可以抑制生物活性。但是,最近有研究表明,柑橘皮与其他材料或高温厌氧发酵下联合厌氧消化更稳定,抑制作用更弱。尽管联合厌氧消化提供了一种经济手段使与某些个别底物有关的难度最小化,但是也要考虑共基质的可用性和可及性。同样地,因为高温发酵的成本相对较高,高温发酵(>55℃)很少被用到。其他的一些方法也已被探索出来,比如微生物的驯化、在AD进程前或者期间去除有毒化学物质。目前,在AD期间抗柠檬烯毒性的方法主要是降低浓度、增加细菌的驯化时间。但是,降低柠檬烯浓度更可取的方法是用物理方法去除AD系统中的柠檬酸,因为同化产生的代谢物具有抑制作用。蒸汽蒸馏现已被证实是一种能从柑橘皮废料中去除多达70%的柠檬烯的方法,但它的能源消耗也很大。除了蒸汽蒸馏和溶剂萃取,吸附被认为是一种去除有机物的方法。例如,Chen et al报道了吸附剂对减少潜在的抑制剂有积极作用,如氨和长链脂肪酸。像沸石、活性炭、膨润土、硅胶这些吸附剂已被报道出可将AD过程中的有毒化学物质除去。虽然生物炭在AD中去除潜在的抑制化学物质的应用还未得到充分的研究,但是有迹象表明生物炭可以吸附单萜化合物。
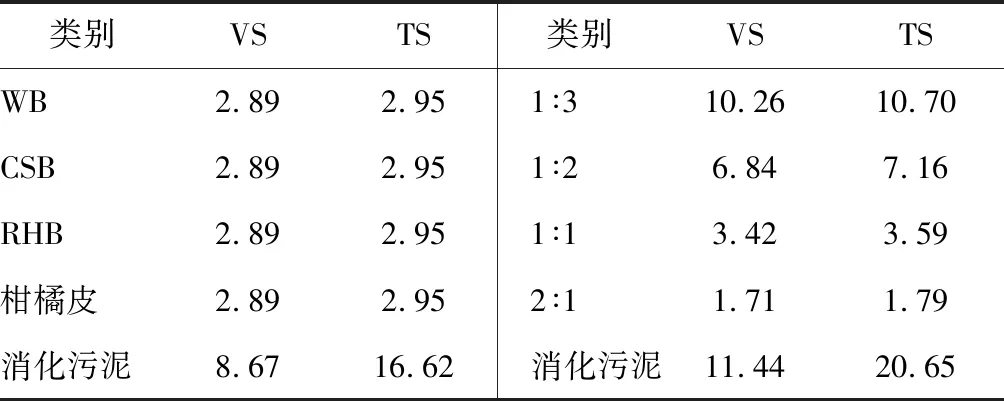
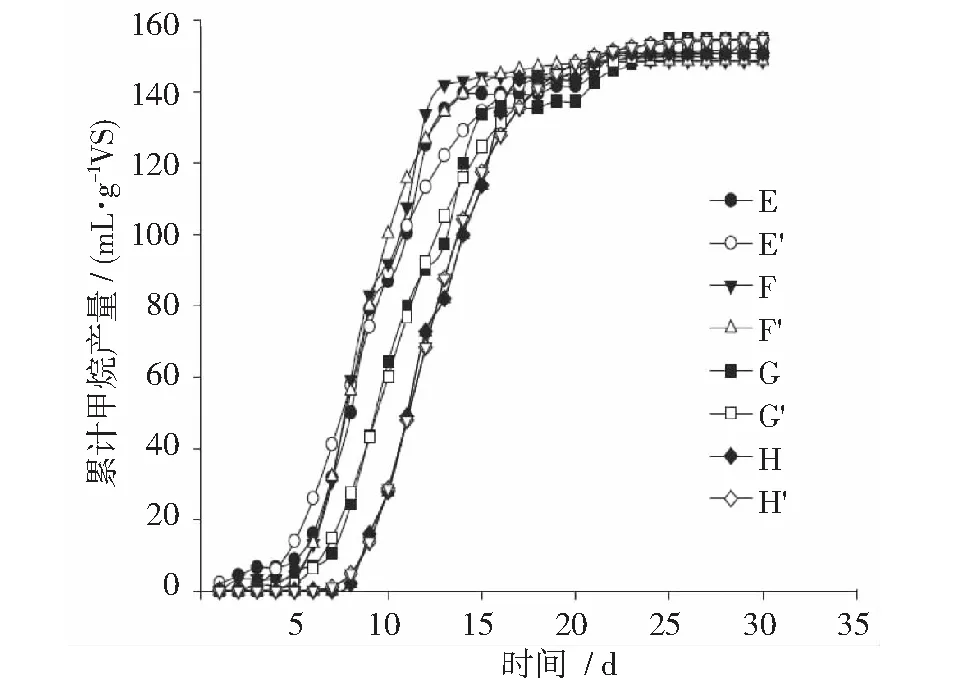
生物炭是由植物衍生生物质在局部或完全无氧环境中进行热处理产生的。热处理通过改变粒子的显微结构来形成一个芳香-脂肪区和一个结晶区,它们由不同内径(ID)的孔隙组成。微孔(ID<2 nm)、大孔(ID>50 nm)、中孔(2 nm 此研究以柑橘皮废料为底物,以消化污泥为微生物接种源。消化污泥由污水处理厂(United Utilities, Preston, UK)提供。对消化污泥进行表征,其所含总固体量为11.0%±0.13%,挥发性固体量为52.1%±0.36%,SCOD含量为27.0±0.23 g·L-1,NH4-N含量为0.71±0.17 g·L-1,pH值为7.26±0.02。柑橘来自英国兰开斯特当地的一家知名超市。清洗后进行榨汁,并称量果皮和果肉重量。柑橘皮废料与32 g柠檬,12 g石灰,53 g橙子,30 g橘子和65 g葡萄皮混合,搅拌均匀并冷冻保存。混合比例根据每年产生的不同柑橘废料的平均数量来选择(FAOSTAT, 2013)。 文中使用了木材生物表(WB),椰壳生物炭(CSB)和稻壳生物炭(RHB)。这些生物炭的制作是在450℃下热解形成。CSB和RHB取自马来西亚,WB取自英国。这些生物炭粒径为1.7~2.0 mm。 柑橘皮废料AD批次实验的研究主要是:1)柑橘皮和生物炭类型; 2)柑橘皮和生物炭比例。控制培养只包括接种、柑橘皮、接种。本研究使用一个500 mL杜兰瓶,带有修改过的橡胶塞,塞上有气体和液体的取样口,气体端口连接一个检测器用于气体体积的测量。在测定甲烷之前,沼气通过一个含有3 mol·L-1NaOH的100 mL杜兰瓶使CO2固定。此外,橡胶塞上还有一个搅拌棒端口,可允许12 V DC电动机在30 rpm下进行机械均化。厌氧反应器在35℃数字水浴锅中培养30 d。根据挥发性固体含量,底物与接种物之比在0.31~0.33之间。由于第一项研究目的是不同类型生物炭的影响,不同的生物炭材料根据总固体干重与柑橘皮以1∶1混合,WB根据总固体干重,柑橘皮和生物炭比例分别为1∶3,1∶2,1∶1。试验方案总结如表1,在500 mL的杜兰瓶内装300 mL的接种物,将其放在35℃的环境下培养2 d,可以显著的减少有机物含量。之后将各组分与底物混合,开始试验。用3 mol·L-1NaOH调节各反应物的pH值大约为7,然后用氮气净化系统1 min去除多余的O2。最后,用1 mm的筛子将沼渣从生物炭中分离出来进一步分析。 通过将样品在105℃的烘箱(Memmert, Germany)和550℃的马弗炉(Carbonite, Sheffield UK)中加热24 h来分析TS和VS含量。分别用于TS和VS的测定(APHA,1998)。测量pH值后,将样品在4500 rpm下离心15 min,上清液用0.45 μm的醋酸纤维素膜过滤,得到可溶性部分。用含有重铬酸盐和二元醇的消化试剂盒测定可溶性组分中SCOD和TFVA,此方法在简单消化后,用定量Hach分光光度法(DR/2800)测定颜色变化。为了测定元素和木质纤维素,需将样品用60℃的烘箱烘干。用元素分析仪将球磨干样进行碳、氮、氢、硫、氧元素的测定。用回流装置和纤维洗洁精进行样品中纤维素、半纤维素和木质素含量的分析;用重量法(ANKOM, USA)测定木质纤维素的组成;复合分离需用超2毛细管柱热解吸GC-MS法测定柠檬烯浓度。先在35℃烘箱中放置2 min,在4℃·min-1时加热至160℃,然后在45 min时加热至300℃,保持10 min。冲洗时,用10 μL注射器吸取5 μL样品,注入吸附树脂、Tenax TA和Carbotrap中,并持续用气吹扫。 表1 柑橘皮发酵、柑橘皮和不同类型生物炭混合发酵的批次实验条件 (g) 2.3.1 阳离子交换容量(CEC) 此分析用1M钠溶液和醋酸铝来测定生物炭的控制和交换能力(Huff etal, 2014)。将乙酸钠和乙酸铝的pH值分别调整至8.2和7,将乙酸钠溶液装入含有4 g生物炭的烧瓶内,置换生物炭表面存在的其他阳离子。经过10 min 500 rpm的离心(Rotana Zentrifugen)后,去掉上清液。此时,生物炭残渣的负极区域覆盖有大量的钠离子。然后将乙酸铵溶液加入生物炭中置换钠离子,将混合物离心,除去上清液,用火焰光度计(JenWay, UK)测量钠离子的位移量。置换钠离子的浓度与生物炭材料的阳离子交换能力成正比。 2.3.2 实际甲烷生产 用带有电子脉冲传感器和数据记录装置的校准提示计对甲烷的产量进行量化。实际甲烷产量是由减去纯接种物的培养液的甲烷产量决定的,尽管在仅有接种物发酵时未观察到可测量的甲烷。总甲烷产量是由生成甲烷的体积除(ACH4)以初始挥发性固体总量(gVS)决定的。 (1) 式中:ACH4为实际甲烷体积;VS为添加的柑橘皮挥发性固体含量。 2.3.3 理论甲烷生产 理论甲烷产量由公式(2)计算得到,用柑橘皮样品中分子式的元素组成,即CaHbOcNdSe。理论甲烷用mL·g-1VS表示。 (2) 2.3.3.1 甲烷转化效率 甲烷转化效率定义为实际甲烷与理论甲烷之比,如公式(3)所示: (3) 2.3.4 修改Gompertz方程 检查所有培养箱的甲烷产量是否符合修改后的Gompertz方程(Zwietering etal, 1990),甲烷产量是细菌生长函数的假设是修改后的Gompertz方程的应用的基础(Zhu etal, 2009)。此模型已应用于确定分批生长中的滞后或者适应阶段(Syaichurrozi)。Gompertz方程如公式(4): (4) 式中:F为累积甲烷产量,mL·g-1VS;A为潜在甲烷产量,mL·g-1VS;r为最大甲烷产量速率,mL·g-1VS·d-1;e为常数,2.718282;λ为滞后期的持续时间;t为累积甲烷产量的时间,d。 2.3.5 数据分析 用Microsoft Excel 2013进行均值、标准差、标准误差的计算,用Sigma plot软件13.0版本用于数据统计分析,经过Shapiro-Wilk正常测试后评定在生物炭类型和其比例之间日常甲烷产量的重要性,Holm-Sidak法用来各组之间平均值的多重比较。显著性检验设定p<0.05。 第1项研究是有关于在30 d的发酵周期下3种不同类型生物炭对柑橘皮厌氧消化的表现。甲烷量在培养第1天就迅速出现,在第1天,WB培养出现最高甲烷产量,对此,CSB和RHB培养具有显著性(p<0.05),因此,需要为代谢中间产物的有机质分解创造一定的条件。但是,1 d之后所有培养下的甲烷生产都趋于稳定到培养3 d后,之后无可测量的甲烷产量。第3天培养的甲烷生产的停滞归因于柑橘皮有机基质中的柠檬烯的存在。已报道出柠檬烯可严重抑制厌氧消化过程和甲烷生成。在第8天的培养中,CSB和WB培养的甲烷产量增加,CSB和WB培养较快的恢复速率表明它们更易吸收柠檬烯。另一方面,RHB培养一直保持抑制状态,直到第13天,然后甲烷产量增加,但是,在培养的第17天观察到甲烷产量显著增加(p<0.05)。RHB培养的甲烷产量最高,此后观察到其甲烷产量速率的连续峰和增加,而CSB和WB培养的甲烷产量持续下降。CSB和WB培养的甲烷产量下降说明可用有机基质已被新陈代谢。Li等报道出活性炭可以吸收可溶性代谢物,例如乙酸,还有可能吸收氢离子,并且生物炭和活性炭在结构上相似。正如预期的一样,只有柑橘皮的培养有最长的抑制期,可持续14 d,这进一步说明生物炭的增加可以提高与柑橘皮一起培养的厌氧菌的恢复速率。这个趋势类似于Watanabe 等得到的结果,他提到粗甘油的厌氧消化中,日本雪松木炭的添加会有更高的甲烷生产活性水平,这是由于柠檬烯对生物炭的吸附作用。此外,Mumme等发现,在厌氧消化系统中生物炭的增加可以减少微生物的滞后期从而加快生长期的开始。但是,柑橘皮厌氧消化中不同类型生物炭的内含物在处理过和未经处理过的生物炭中的总甲烷产量都没有明显增加(p>0.05)。CSB,WB,RHB和柑橘皮培养的累计甲烷产量分别在第9天,23天,20天完成,同样地,Mumme等在氨培养的缓解研究中检测到改性生物炭有更高的累计甲烷产量。从柑橘中获得的甲烷产量与Serrano等的结果相似,他记录了在每0.4~1.6 gVS·L-1d-1的OLR是165 mL·g-1VS。高甲烷转换率表明这个厌氧消化过程是有效的,因此CSB培养的生物炭类型是最有效的处理。 第2项研究是观察柑橘皮厌氧消化中不同生物炭比例在查过30 d培养周期下的表现。此研究用四种不同柑橘皮与WB的混合比例,即1∶3,1∶2,1∶1,2∶1,测定他们的速率和累计甲烷产量。累计甲烷产量在第1天培养时迅速开始,2∶1的培养有最低甲烷产量速率(p>0.05)。但是,培养3 d后,除了1∶3培养外,其它培养的甲烷产量达到一个无法测量的速率的平衡,平均值在2.69±1.09 mLCH4·g-1VS·d-1,在第9天时增加到7.37±3.01 mLCH4·g-1VS·d-1。如前面提到的,柑橘皮中柠檬烯的含量被认为具有明显抑制甲烷生成和减少甲烷产量的作用,结果表明,甲烷生成的抑制作用随柑橘皮和WB的比例的降低而减弱,1∶3,1∶2,1∶1培养分别抑制1 d,2 d,3 d,而2∶1培养会抑制5 d,这支持了WB减少柠檬烯生物活性的观点,这个趋势与吸附剂浓度增加对山梨酸盐的去除有积极作用的结果一致。在只有柑橘皮培养中的抑制率可持续7 d,含有WB培养的为1~5 d,而且相对于只有柑橘皮培养的总甲烷产量时间即20 d,WB的增加会减少其总甲烷产量时间,平均为17 d。生物炭被认为是可以提供一个大的比表面积,可以让微生物细胞增殖。据报道,微生物细胞可减小共生细菌和产甲烷菌的距离,这增加了挥发性脂肪酸的氧化量以及产氢量。但是,WB比例的增加并没有增加测试培养的总甲烷产量,因为它并没有明显放的变化(p>0.05)。因此,我们可以得出结论,即生物炭的增加提高了柑橘皮厌氧消化中的甲烷回收率。但是比较不同生物炭和不同比例的效果时这个参数不能作为一个很好的参考,这是因为它们的总甲烷产量相对来说是相似的。 甲烷产量的程度与产甲烷种群的增长有直接的关系。修改的Gompertz方程拟合了累积甲烷生产曲线和获得的参数值(见表2)。得到的回归≥0.990。修改的Gompertz方程拟合了柑橘皮曲线,如图1~图2所示是柑橘皮生物炭类型和比例,数据表明生物炭的增加导致滞后期长度发生变化,它与生物炭的吸附特性和柠檬烯生物活性的降低有关,从而提高了微生物细胞的回收率。柠檬烯和生物炭具有疏水性,所以范德华力将芳香烃吸附到炭质吸附剂上。RHB培养从第3天~13天的长滞后期和不可测量的甲烷产量速率说明了柠檬烯的抑制作用。 A-CSB和柑橘皮混合发酵实验;A′-CSB和柑橘皮混合发酵模型;B-WB和柑橘皮混合发酵实验;B′- CSB和柑橘皮混合发酵模型;C-RHB和柑橘皮混合发酵模型;C′ -RHB和柑橘皮混合发酵模型;D-仅柑橘皮混合发酵实验;D′-仅柑橘皮混合发酵模型。图1 不同发酵原料的累计甲烷产量 E-WB和柑橘皮1∶3实验; E′-WB和柑橘皮1∶3模型; F-WB和柑橘皮1∶2实验; F′-WB和柑橘皮1∶2模型; G-WB和柑橘皮1∶1实验; G′-WB和柑橘皮1∶1模型; H-WB和柑橘皮2∶1实验; H′-WB和柑橘皮2∶1模型.图2 不同发酵原料的累计甲烷产量 RHB有最高的去除率,为88.82%,高于其它生物炭材料。生物炭比例方面,柠檬烯的剩余量会随着生物炭在每个培养中的增加而减少,1∶3培养有最高的去除率,为95.12% ,高于其它培养。柠檬烯浓度的降低与吸附到生物炭上以及生物降解有关,还有可能是因为消化污泥的加入导致其稀释。只有柑橘皮的控制培养记录了比较生物炭比例时柠檬烯残渣的第二低浓度和比较生物炭类型时的最高浓度,含柑橘皮培养的值是10.48 mg·L-1,去除率是93.86% ,由于没有生物炭材料的添加,预计控制培养会有最高的柠檬烯残渣浓度,此现象只能归因于柠檬烯的生物降解。有迹象表明,柠檬烯化合物在AD期间可以被转换成其它代谢产物。柑橘皮培养有最低甲烷产量和最长滞后期,说明柠檬烯或假定代谢物抑制甲烷生成 表2 拟合累积甲烷生产曲线的参数值 用扫描电镜法对消化污泥和柑橘皮废料接种前后不同生物炭的微生物形态作比较。研究表明,SEM图像显示生物炭材料上有微生物的存在,除了CSB外,微生物成功的移植在生物炭上,生物炭表面的整体粒度为微生物细胞或者可能是生物膜提供了有利的环境。WB上的形态数量和多样性最大,这可能是由于材料表面有丰富的大孔隙,但是需要更详细的生化和分子生物测试去证实这一定论。奇怪的是,CSB培养未表现出明显的微生物细胞附着,可能是细胞在样品准备时被清除。生物炭上群体的出现表明是微生物细胞的固定化作用,支持产甲烷菌生成。 十分明确在污染土壤和沉积物只加入炭黑,包括生物炭可以减少有毒化学物质的移动性和生物活性。据作者所知,生物炭对厌氧消化和甲烷生成的影响还未被报道。本研究首次探讨了生物炭对厌氧消化中柠檬烯的甲烷生产的影响,研究表明,在比较不同生物炭对柑橘皮厌氧消化的影响时,WB培养有最短滞后期,而CSB培养有最高甲烷产量。研究还表明柑橘皮与生物炭比例的增加会延长微生物的滞后期,这说明WB和CSB材料在较高比例时对维持厌氧消化进程的稳定性是有效的,尤其是在底物诱导抑制期。 (农业部沼气科学研究所 张冀川 译自:Impact of biochar on the anaerobic digestion of citrus peel waste[J].Bioresource Technology, 2016,(216):142-149)2 材料与方法
2.1 底物与接种
2.2 实验设计与步骤
2.3 化学分析

3 结论与讨论
3.1 不同生物炭对柑橘皮厌氧消化的影响
3.2 不同生物炭比例对柑橘皮厌氧消化的影响
3.3 动力学因素

3.4 柠檬烯去除效率

3.5 生物炭的形态变化
4 结论
