饲料和饲料原料等25 种样品中16 种真菌毒素超高效液相色谱-三重四级杆/线性离子阱串联质谱法基质效应的研究
2019-12-04符金华杨琳芬董泽民徐国茂姜文娟刘国花周仁丹
符金华, 杨琳芬, 董泽民 , 徐国茂, 姜文娟, 叶 金,吴 宇, 邢 磊, 刘国花, 樊 晶, 周仁丹, 廖 丰
(1.江西省兽药饲料监察所,江西南昌330029;2.国家粮食局科学研究院,北京100037;3.南昌大学分析测试中心,江西南昌330047;4.双胞胎(集团)股份有限公司,江西南昌330096)
随着检测技术的飞速发展, 仪器设备的迭代更新,高精密度的仪器应用越来越普遍。 其中,液相色谱与质谱联用技术(LC-MS/MS)是目前痕量物质定性和定量分析的首选, 其兼具液相色谱与质谱技术的优点,具有高灵敏度和精确度,并大大缩 短 分 析 时 间 (Ito 等,2002; Matuszewski 等,2000;Sutherland 等,2000;Fu 等,1998)。 然而,在含相同浓度的待测物标准溶液和饲料等畜产品样品通过LC-MS 分析时,某些待测物的质谱响应发生显著的增强或抑制, 尤其在缩短色谱分离时间时更为显著(Bogialli 等,2003),这种现象即为质谱检测中的“基质效应”(ME)。基质效应产生的主要原因有以下几个方面:(1)样品中其他基质成分与目标物在离子化阶段发生互相竞争, 从而会显著地降低或增加目标物的离子化效率及离子强度;(2)源于样品在前处理过程中使用的部分试剂或药物溶剂,如吐温-80、聚乙二醇-400(Xu 等,2010)、Tris 缓冲液、NADPH 等(Pellati 等,2002);(3) 不恰当的内标会与待测物竞争液滴表面的电荷而导致离子强度变化(Liang 等,2010)。因此,基质效应严重影响目标化合物的LC-MS 定量分析准确度和精密度, 在应用LC-MS 技术测定时,必须重视基质效应的影响及作为方法学考察的重要部分(林涛等,2015; 卞愧等,2014;Rogatsky 等,2005;Ito 等,2002)。
真菌毒素具有高毒性和强致癌性, 严重危害人、畜健康,威胁食品安全并造成巨大的经济损失(符金华等,2017)。 本研究针对目前农产品、饲料、饲料原料中真菌毒素污染较为普遍的现状,旨在利用超高效液相色谱-三重四级杆/线性离子阱串联质谱(Qtrap-UPLC-MS/MS) 技术建立一种快速定性、定量分析饲料、饲料原料、奶和肉样品中16 种真菌毒素的分析方法(符金华等,2019、2017;张大伟等,2018;杨琳芬等,2017)。在整个定量线性范围内,以目标物在空白基质液中的峰面积与溶剂中峰面积的百分比来系统地探究常见的5 大类配合饲料(鱼、猪、鸭、鸡和牛)、5 种饲料原料(豆粕、鱼粉、菜籽粕、菜粕及DDGS)等25 种样品的基质效应对常见16 种真菌毒素测定的干扰, 并提出针对性的消除措施, 为提高Qtrap-UPLC-MS/MS 法测定16种真菌毒素结果的准确度和精密度提供数据支持。
1 材料和方法
1.1 仪器和试剂 质谱型号为Sciex4500 Qtrap(美国Sciex 公司);液相为岛津LC-20AD XR(日本Shimadzu 公司);赫西离心机(中国湖南赫西仪器装备有限公司);HY-3 多功能振荡器 (中国江苏光都机电设备有限公司):Milli-Q 超纯水纯化系统 (美国Millipore 公司),METTLER TOLEDO ME104 分析天平(中国梅特勒-托利多(上海)有限公司)。
雪腐镰刀菌烯醇(NIV)、脱氧雪腐镰刀菌烯醇(DON)、3-乙酰基脱氧雪腐镰刀菌烯醇(3-Ac-DON)、15-乙酰基脱氧雪腐镰刀菌烯醇 (15-Ac-DON)、脱氧雪腐镰刀菌烯醇-3-葡萄糖苷(DON-3G)、T-2 毒素(T-2)、HT-2 毒素(HT-2)、玉米赤霉烯酮(ZEN)、赭曲霉毒素A(OTA)、伏马毒素B1(FB1)、伏马毒素B2(FB2)、杂色曲霉毒素(ST)标准溶液 (浓度为0.2 ~ 20 μg/mL,Romer 公司);黄曲霉毒素B1(AFB1)、黄曲霉毒素B2(AFB2)、黄曲霉毒素G1(AFG1)、黄曲霉毒素G2(AFG2)标准溶液(浓度为0.03 ~1 μg/mL,Sigma-Aldrich 公司);13C 标记的黄曲霉毒素(B1、B2、G1、G2)、脱氧雪腐镰刀菌烯醇、 雪腐镰刀菌烯醇、3-乙酰基脱氧雪腐镰刀菌烯醇、玉米赤霉烯酮、赭曲霉毒素A、伏马毒素(B1、B2)、T-2 毒素、HT-2 毒素、杂色曲霉毒素的14 种稳定同位素标准溶液 (浓度为0.01 ~2.5 μg/mL,Romer 公司);甲醇、乙 腈(HPLC 级,Fisher 公司);乙酸铵、甲酸、乙酸(HPLC 级,美国Sigma 公司);0.2 μm PTFE 膜针头过滤器(PALL公司)。
1.2 液相色谱条件 Waters 公司CORTECSTM UPLC C18柱(100 mm × 2.1 mm,1.6 μm);柱温40℃; 进样量2 μL; 流动相A 为甲醇,B 为含0.1%(体积分数)的甲酸和1 mmol/L 乙酸铵的水溶液;流速为0.3 mL/min;采用梯度洗脱。
1.3 质谱条件 加热电喷雾离子源温度为500 ℃;喷雾电压为5500 V; 离子传输管温度为320 ℃;gas1 和gas2 均为50 psi,气帘气为35 psi。 EPI 扫描速度10000 Da/s, 扫描质量范围为50 ~1000,分段多反应监测扫描模式,MRM 检测窗口设置为30 s,锥孔电压50 V;正、负离子采集。
1.4 标准溶液配制 分别移取一定体积的16 种霉菌毒素标准溶液于10 mL 容量瓶中,用蒸馏水定容, 得到16 种霉菌毒素的混合标准储备液,于-20 ℃保存,浓度详见表1。

表1 真菌毒素混合标准储备液及相应的稳定同位素混合溶液的浓度 g/mL
稳定同位素内标混合工作液: 根据表1 相应的稳定同位素混合溶液的浓度, 分别移取一定体积的14 种霉菌毒素稳定同位素单标溶液于4 mL储液瓶中, 用水稀释至2 mL 配制稳定同位素混合标准溶液,充分混匀后于-20 ℃避光保存。 准确移取混合标准中间液适量,用乙腈-水-乙酸溶液(35:64.5:0.5)逐级稀释,配制成不同浓度系列的混合标准工作液。 向400 μL 内插管中加入20 μL 14 种稳定同位素混合工作液, 再分别吸取180 μL系列标准工作液于内插管中, 涡旋混匀后准备上机检测。
1.5 样品前处理方法 称取饲料样品 (5±0.1)g于50 mL 离心管中,准确加入20 mL 乙腈-水-乙酸溶液(V/V/V,70:29:1,)提取溶剂,涡旋2 min,振荡30 min,以4000 r/min 离心10 min 使固液分离。 准确转移0.5 mL 上清液于1.5 mL 离心管中,加入0.5 mL 水稀释, 涡旋混匀1 min, 然后以12000 r/min 离心10 min,取上清液用0.22 μm 的PTFE 滤膜过滤, 吸取20 μL 预先涡旋混匀的稳定同位素混合溶液于400 μL 内插管中, 再加入180 μL 的样品滤液,混合后待测。
2 结果与讨论
本研究以目标物在空白基质液中的峰面积与溶剂中峰面积的百分比来系统评估25 种样品的基质效应。 当结果接近100%时,表明无明显的基质效应,而高于100%说明有基质增强效应,低于100%则说明有基质抑制效应。16 种真菌毒素在5大类配合饲料和5 种饲料原料中的基质效应如图1 ~图6 所示。
由图1 可知,16 种真菌毒素在鱼饲料中的基质效应为48.6% ~145.4%。其中T-2/OTA/FB1/FB2四种毒素的基质效应为101.7% ~145.4%, 且有不同程度依次增大的基质增强效应。 其他毒素都存在不同程度的基质抑制效应, 基质效应为48.6% ~99.5%。 同时,五种鱼配合饲料的基质效应呈现一致规律性变化,16 种真菌毒素的整体基质效应较强, 这可能与鱼饲料中含有较多的植物性原料有关, 如菜粕和菜籽粕会引入大量色素等有机质, 样品的提取上清液颜色明显比其他基质的更深,最终导致基质效应较强。
图2 结果显示,16 种真菌毒素在猪饲料中的基质效应为41.8% ~146.4%, 其中ST/T-2/OTA/HT2/FB2/FB1六种毒素的基质效应为101.8% ~146.4%,且为依次增强的基质增强效应。 其他的毒素基质效应为41.8% ~99.6%, 都存在不同程度的基质抑制效应, 且6 种猪饲料的基质效应也呈现一致规律性变化。 同时, 绝大部分毒素的基质效应处于61.5% ~109.5%, 相对鱼饲料来说,其基质效应的程度更弱。
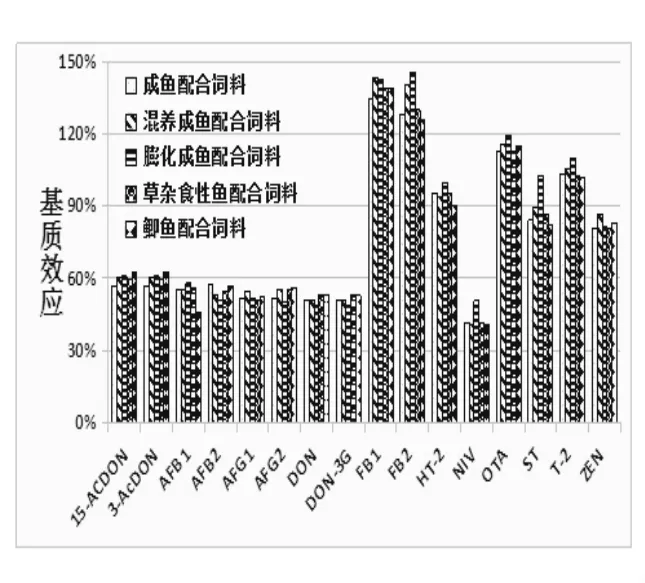
图1 16 种真菌毒素在5 种鱼饲料基质中的基质效应

图2 16 种真菌毒素在6 种猪饲料基质中的基质效应
由图3 所示,16 种真菌毒素在鸭饲料中的基质效应为43.2% ~148.1%, 其中肉鸭料中T-2/OTA/FB2/FB1四种真菌毒素的基质效应为101.7% ~148.1%,呈现基质增强效应。其他的毒素都存在不同强弱的基质抑制效应,基质效应为70.6% ~98.6%。然而,蛋鸭饲料出现了不同于另两种鸭饲料的基质效应,DON/3-AcDON/15-Ac-DON 三种真菌毒素出现了106.8% ~137.4%的基质增强效应,而出现在鱼、猪等饲料中基质增强的T-2/OTA/FB2/FB1等毒素出现基质抑制现象,这个可能与蛋鸭料中的蛋白质等其他营养成分含量较高有关。

图3 16 种真菌毒素在3 种鸭饲料基质中的基质效应
由图4 可知,16 种真菌毒素在不同种类鸡饲料中的基质效应为42.1% ~148.3%。 其中T-2/OTA/HT-2/FB2/FB1五种毒素出现了依次增大的基质增强效应, 基质效应为103.9% ~148.3%,FB2/FB1两种毒素的基质增强效应严重。其他的毒素都存在不同强弱的基质抑制效应, 且绝大部分毒素的基质效应处于67.5% ~89.1%, 说明该基质效应抑制程度较其他饲料要弱一些。 同时,五种鸡饲料的基质效应呈现一致规律性变化。

图4 16 种真菌毒素在5 种鸡饲料基质中的基质效应
由图5 可知,16 种真菌毒素在犊牛饲料中的基质效应为50.3% ~140.8%,其中ST/T-2/HT-2/OTA/FB2/FB1六种毒素出现了依次增大的基质增强效应, 基质效应为105.4% ~140.8%, 且FB2/FB1两种毒素的基质增强效应严重。 其他的毒素都存在不同强弱的基质抑制效应, 且绝大部分毒素的基质效应处于66.0% ~86.7%, 说明该基质抑制效应程度不太明显。
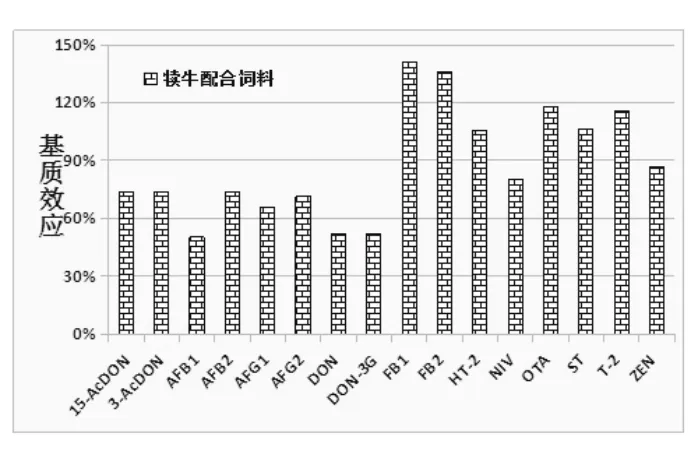
图5 16 种真菌毒素在牛饲料基质中的基质效应
由图6 可知,16 种真菌毒素在5 种常见饲料原料中的基质效应为30.7% ~148.6%,其中T-2/OTA/FB2/FB1四种毒素出现了依次增大的基质增强效应,基质效应为100.6% ~148.6%,其他的毒素都存在不同强弱的基质抑制效应。 豆粕和鱼粉的基质效应较弱,但菜粕、菜籽粕和DDGS 三种原料的基质效应严重,且DDGS 最为严重。这可能是由于菜粕中含有较多的色素, 菜籽粕中含有较多的色素和油脂,DDGS 中含有大量的小分子蛋白质和有机酸等有机物, 从而产生了较强的基质干扰。同时,五种饲料原料的基质效应呈现一致规律性变化。
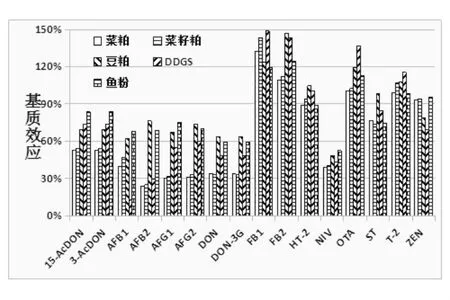
图6 16 种真菌毒素在饲料原料基质中的基质效应
以上研究结果表明, 在这几大类常见动物饲料和原料的基质中存在一定的基质增强或抑制效应。T-2/OTA/FB2/FB1等四种真菌毒素的基质增强效应明显,AFB1/AFB2/DON/DON-3G/NIV 等几种毒素基质抑制效应明显。因此,要精确定量测定饲料及原料中常见的16 种真菌毒素,需要采取正确的措施来尽量消除或减弱基质效应。 基质匹配标准曲线是一种常见的减少基质效应的方法, 但是由于自然状态下真菌毒素的污染具有普遍性,导致实际工作中不易获得均不含16 种真菌毒素的空白基质样品, 特别是完全匹配的基质样品更是难上加难, 从而使得代表性基质样品的普适性面临挑战,具有较大的不确定性,最终导致基质匹配定量在标准化检测中的应用受到局限。然而,稳定同位素法是另一种能够较好消除基质干扰的途径。为了更好消除基质效应的影响,本实验采用稳定同位素稀释法进行定量, 以确保结果的准确可靠。结果证明,稳定同位素稀释法能够精确定量测定饲料及原料中常见16 种真菌毒素。
以稳定同位素稀释法进行定量结果表明,在一定的线性范围内,16 种真菌毒素线性关系良好,相关系数(r2)均>0.998。 对混合标准添加溶液进行逐级稀释,以3 倍信噪比(S/N)计算检出限(LOD) 为0.12 ~65 μg/kg,10 倍S/N 计算定量下限(LOQ) 为0.50 ~200 μg/kg。 16 种真菌毒的LOQ 均低于我国和欧盟规定的饲料及原料中的限量值,取16 种霉菌毒素含量较低(低于方法检出限) 的5 类常见配合饲料和5 种常见饲料原料样品。 分别添加高、中、低3 个浓度水平的混合标准溶液,每个加标水平进行6 次重复实验。 16 种霉菌毒素在5 种常见配合饲料中的回收率绝大数都在70.2% ~129.5%,16 种霉菌毒素在5 种常见配合饲料中的回收率绝大数都在66.4% ~128.9%, 且16 种真菌毒素回收率的RSD 值为0.1% ~14.5%。 两大类基质中的回收率和RSD 均符合相关法规的检测要求, 说明本方法准确度和精密度可以满足日常检测的需要。
3 结论
本试验结果表明,5 大类配合饲料和5 种饲料原料等25 种样品基质对16 种真菌毒素的精确定量测定均存在不同程度的基质增强或抑制效应。 采用同位素稀释技术能够有效地消除基质效应, 方法准确度和精密度可满足于饲料及饲料原料中常见真菌毒素的残留检测分析的需要, 为快速精准测定饲料及原料中16 种真菌毒素的污染防控提供了科学参考。
