不同光照节律对内毒素血症大鼠的骨骼肌代谢的影响
2019-11-29程旻桦李孝尧虞文魁
陈 鸣,陈 妍,程旻桦,李孝尧,虞文魁
0 引 言
脓毒症是重症医学科中常见的危重症。此类患者病情复杂,常合并多个器官功能障碍[1],治疗费用高昂,加速康复对其重要性自然不言而喻。但由于所需的医疗操作与监测频繁,其在重症医学科治疗过程大多暴露在异常光照节律中[2]。光照是昼夜节律调节的重要外界因素,是生物节律调节中的重要环节,紊乱的光周期可能引起正常机体一系列神经内分泌及代谢改变,从而导致机体的高分解代谢,特别是骨骼肌萎缩[3],不利于患者加速康复。考虑到脓毒症亦是以蛋白质过度消耗为主的异常高分解代谢为特征[4],因此探索脓毒症的分解代谢与光照节律的关系及机制对于加速病情康复有一定意义,但此类的研究相对缺乏。下丘脑弓状核是重要的能量调节中枢,弓状核的POMC 神经元已被证实在内毒素血症大鼠的骨骼肌消耗中发挥重要作用[5]。下丘脑视交叉上核是重要的生物节律调节中枢,过度光照能够通过干预视交叉上核的生物时钟反馈环,破坏机体的节律同步性,引起机体节律紊乱和代谢改变[6],因此下丘脑可能是光照节律影响能量代谢的枢纽。本研究旨在比较不同的光照节律下脂多糖(lipopolysaccharide,LPS)造模的内毒素血症大鼠的骨骼肌消耗,以及下丘脑水平的相关改变,来研究光照节律对脓毒症状态下机体的代谢影响和机制,探寻能降低骨骼肌分解代谢和消耗的最优化光照节律,以加速脓毒症患者的康复。
1 材料与方法
1.1 实验动物及饲养条件健康清洁级成年雄性SD 大鼠54 只,体重(220±20)g,由南京大学医学院附属金陵医院比较医学科提供。实验动物合格证号:20130016003781,饲养环境:温度稳定在24℃,相对湿度为45%,保持良好的通风条件,使用标准实验动物饲料和饮用水喂养,动物自由进食饮水。实验干预前给予12h/12h光照-黑暗的自然光周期。
1.2 动物模型建立及分组适应环境1 周后,将54只大鼠随机数字表法分为3组,对照组、规律光照组和持续光照组,每组18只。规律光照组和持续光照组大鼠给予腹腔内注射LPS(第1 天5 mg/kg,随后6 d 2 mg/kg),构建内毒素血症模型;对照组大鼠腹腔注射等体积的等渗盐水作为对照。所有动物均单笼饲养,暴露于200 lux光强度的白炽灯照射下持续7 d。其中,对照组和规律光照组的大鼠保持12 h/12 h 光照-黑暗循环,即上午8:00 开灯,晚上8:00关灯;持续光照组大鼠保持7 d 不间断的持续光照条件。
1.3 组织标本获取与保存每隔24 小时记录大鼠的体重变化和食物摄入量。第8 天时,为检测钟基因节律分别在08:00 及其后每隔4 小时的时间点上,在每组中随机选择3 只大鼠进行处死。腹腔注射超量的2%戊巴比妥钠(0.5 mL/100 g)麻醉大鼠,并用200 mL等渗盐水经心脏灌注后,取下双侧趾长伸肌(EDL)称重,放入2 mL 冻存管,冻存在液氮罐中;取大鼠下丘脑,同样冻存于液氮罐中,以待检测。若取材时间点处于光照-黑暗周期中的黑暗阶段,进行所有操作时需在红光灯下进行。
1.4 骨骼肌代谢产物及萎缩基因表达水平检测采用高效液相色谱-质谱联合法检测3-甲基组氨酸(3-MH)与酪氨酸水平。从液氮中取出大鼠趾长伸肌解冻后,称取约0.2 g 样本加入1.00 mL 预冷的30%乙醇水,振荡研磨后,离心获取上清液。利用3-MH 和酪氨酸的标准品得到标准曲线,分别取各待测样本液进行上机检测,记录各样本峰浓度下面积。根据所得标准曲线计算各样本中3-甲基组氨酸和酪氨酸的含量。
采用实时荧光定量PCR 检测骨骼肌萎缩基因MAFBx 和MuRF-1 的表达水平。取100mg 肌肉组织加入液氮研磨成粉末,使用总RNA 提取试剂盒进行RNA 提取,使用cDNA 合成试剂盒(Thermo,USA)通过逆转录合成cDNA。用于RT-PCR 分析的引物序列设计为:MAFBx:上游5'-GTCTCACGATCACCGACCTG-3',下 游5'-CTTCTTGGGTAACATCGCACA-3';MuRF-1:上游5'-GTGCCTACTTGCTCCTTGTGC-3',下游5'-GGCGTAGAGGGCGTCAAACT-3'。
1.5 炎症因子水平检测采用实时荧光定量PCR检 测。 引 物 序 列 为:TNF- α:上 游5'-CCACGCTCTTCTGTCTACTG-3', 下 游 5' -GCTACGGGCTTGTCACTC-3';IL-1β:上游5'-TTCAAATCTCACAGCAGCAT-3', 下 游 5'-AGGTCGTCATCATCCCAC-3'。
1.6 钟基因BMAL1 和CLOCK 的mRNA 水平检测采用实时荧光定量PCR 检测BMAL1 和CLOCK的表达水平。取100 mg 下丘脑组织加入液氮研磨成粉末,使用总RNA 提取试剂盒进行RNA 提取,使用cDNA 合成试剂盒(Thermo,USA)通过逆转录合成cDNA。用于RT-PCR 分析的引物序列设计为:BMAL1:上 游5'-TTCCCACAGCTTGCAGCTT-3',下游5'-TTTTGGGCCGCCTTCTC-3';CLOCK:上游5'-TCACCACGTTCACTCAGGACA-3', 下 游 5'-AAGGATTCCCATGGAGCAA-3'。
1.7 POMC mRNA及蛋白表达采用实时荧光定量PCR 检测POMC mRNA,引物序列为:上游5'-ACCTCACCACGGAAAGCAAC-3',下 游5'-TCAGTCAAGGGCTGTTCATCTC-3'。采用免疫印迹检测POMC蛋白表达。取下丘脑组织在液氮下研磨,使用裂解液裂解,通过SDS/PAGE分离,用于Western blot印迹分析。
1.8 统计学分析采用SPSS 24.0 软件进行统计分析,定量资料均采用均数±标准差(xˉ±s)表示。所有组间检验使用单因素方差分析,两组间比较使用Tukey 检验。以P ≤0.05 为差异具有统计学意义。使用Matlab 软件进行余弦分析评估BMAL1 和CLOCK 表达的节律性,选择公式F(t)= M + A * cos(t*π/12+φ),对6 个时间点上的基因表达量进行正弦函数拟合得到M,A 和φ 的值。余弦分析根据SE(A)/A 评估准确性,当SE(A)/A <0.3 时确定存在昼夜节律性。
2 结 果
2.1 大鼠摄食、体重、骨骼肌消耗和下丘脑炎症因子表达持续光照组的摄食量、体重增长比和趾长伸肌/体重比明显低于规律光照组(P <0.05),且两组均明显低于对照组(P <0.01)。持续光照组骨骼肌代谢产物3-MH 和酪氨酸以及萎缩基因MAFbx 和MuRF-1 mRNA 以及下丘脑中TNF-α和IL-1β mRNA的表达水平较规律光照组显著升高(P <0.05),且两组均明显高于对照组(P<0.05)。见表1。
2.2 持续光照影响下丘脑生物钟基因的节律性表达处于正常光照周期的对照组和规律光照组大鼠,其钟基因BMAL1和CLOCK的表达均具有明显节律性[SE(A)/A<0.3]。BMAL1 的表达在光照期开始时最高,而CLOCK 在光照期间表达水平高,在黑暗期表达降低。在持续光照组中,BMAL1和CLOCK的表达均失去了节律性(SE(A)/A>0.3,见图1。
2.3 POMC mRNA在下丘脑中的表达持续光照组下丘脑中POMC mRNA 的表达较规律光照组明显提高(P<0.05),且两组均高于对照组(P<0.01)。见图2。
表1 光照节律对内毒素血症大鼠相关指标表达的影响(±s)Table1 Effects of light rhythm on food intake,body weight,skeletal muscle wasting and hypothalamic inflammatory factor expression in endotoxemia rats(xˉ±s)
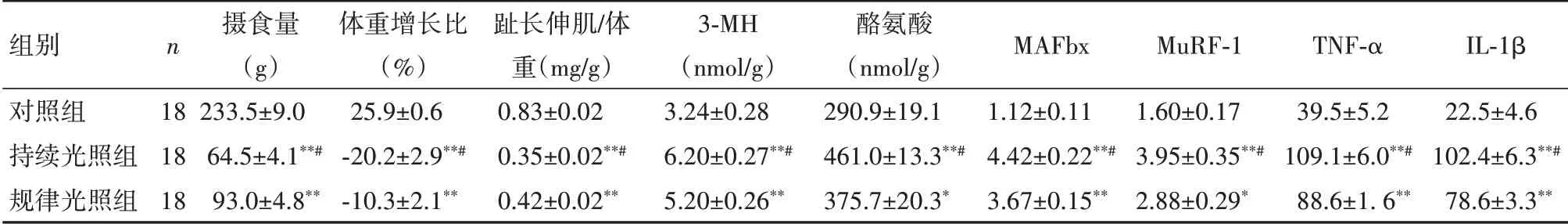
表1 光照节律对内毒素血症大鼠相关指标表达的影响(±s)Table1 Effects of light rhythm on food intake,body weight,skeletal muscle wasting and hypothalamic inflammatory factor expression in endotoxemia rats(xˉ±s)
与对照组相比,*P <0.05,**P <0.01;与规律光照组相比,#P <0.05
组别对照组持续光照组规律光照组n 18 18 18摄食量(g)233.5±9.0 64.5±4.1**#93.0±4.8**体重增长比(%)25.9±0.6-20.2±2.9**#-10.3±2.1**趾长伸肌/体重(mg/g)0.83±0.02 0.35±0.02**#0.42±0.02**3-MH(nmol/g)3.24±0.28 6.20±0.27**#5.20±0.26**酪氨酸(nmol/g)290.9±19.1 461.0±13.3**#375.7±20.3*MAFbx 1.12±0.11 4.42±0.22**#3.67±0.15**MuRF-1 1.60±0.17 3.95±0.35**#2.88±0.29*TNF-α 39.5±5.2 109.1±6.0**#88.6±1.6**IL-1β 22.5±4.6 102.4±6.3**#78.6±3.3**

图1 内毒素血症大鼠下丘脑生物钟基因的昼夜表达水平Figure 1 Diurnal expression level of circadian clock gene in hypothalamus in rats with endotoxemia
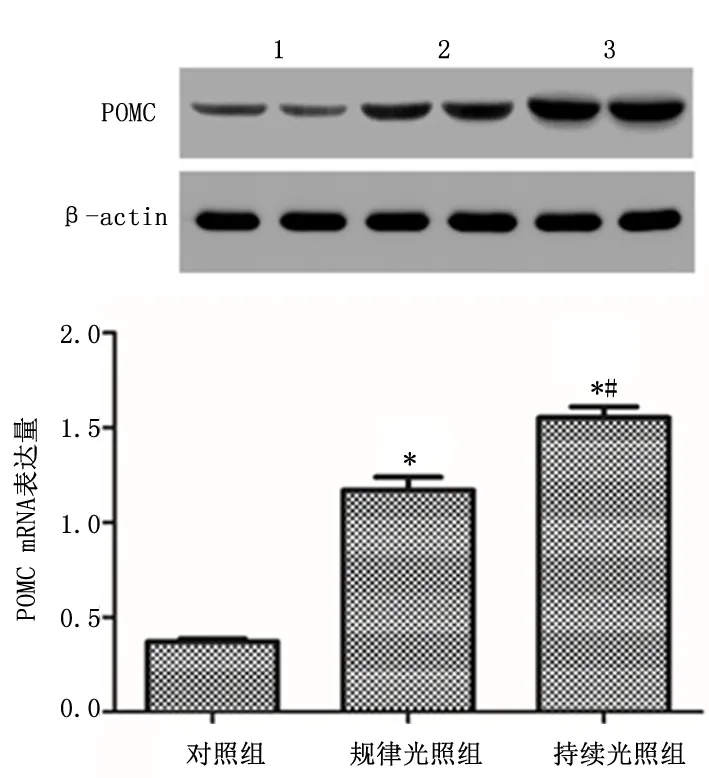
图2 持续光照对内毒素血症大鼠下丘脑POMC 表达的影响Figure 2 Effects of continuous illumination on POMC expression in hypothalamus of endotoxemia rats
3 讨 论
生物节律、睡眠与摄食是机体能量调节的重要组成部分[7],节律异常能够引起弓状核摄食/厌食神经元表达异常[8]。因密切监护与强化治疗的需要,持续光照在ICU十分常见。目前已知持续光照能够在正常机体引起摄食、神经内分泌的改变和高分解能量代谢[9-10]。亦有研究表明脓毒症患者的高分解代谢和由此引起的骨骼肌萎缩常常导致更多的并发症和康复的延迟,甚至不良预后[11],但脓毒症状态下的光照对骨骼肌的影响及其机制国内外报道较少。本研究结果表明,与持续光照相比,暴露于正常规律光照周期的内毒素血症大鼠的肌肉萎缩基因MuRF-1 和MAFbx 表达下调,骨骼肌代谢产物3-MH 和酪氨酸的水平显著下降,光照对内毒素血症大鼠代谢与摄食的作用明显强于健康大鼠[12],证实了正常规律光照对脓毒症状态下减少骨骼肌消耗的重要意义。
我们的前期研究证实在LPS诱导的内毒素血症大鼠中,POMC 神经肽在下丘脑中的高表达与内毒素血症大鼠的肌肉萎缩具有显著相关性,抑制POMC 神经元表达能够显著缓解LPS 诱导下的肌肉萎缩[13]。而下丘脑视交叉上核是哺乳动物的昼夜节律调节中枢,光照则是中枢节律的主要调节因素。视网膜感光细胞接受光照刺激,并将信息传递至视交叉上核,继而向下丘脑的其他核团发送信号,调节外周组织的昼夜节律,最终实现机体生物节律的同步化[14]。保持昼夜节律的重要分子基础是存在于大多数细胞中的生物节律反馈环。该反馈环主要由钟基因BMAL1、CLOCK、CRY 和PER 等组成,其中基因BMAL1 和CLOCK 的蛋白产物组成二聚体,为反馈环提供基本驱动力,发挥正向调节作用[15]。一系列研究表明,基因BMAL1 和CLOCK的mRNA 在12h/12h光照-黑暗循环中的丰度分布表现出节律性,其在视交叉上核区域的表达存在峰值[16]。当给予持续光照刺激时,BMAL1 和CLOCK基因的表达节律出现异常[17]。本研究进一步观察了内毒素血症大鼠接受不同的光照刺激对下丘脑BMAL1 和CLOCK 基因的表达节律和POMC 神经肽活化的影响,发现在LPS诱导的内毒素血症大鼠中,持续光照刺激能够破坏下丘脑BMAL1 和CLOCK 基因的表达节律,显著提高POMC 神经肽的表达。提示持续光照刺激引起钟基因BMAL1 和CLOCK 的表达节律紊乱,继发下丘脑弓状核POMC 神经肽的异常高表达是其导致内毒素血症大鼠骨骼肌消耗的可能机制之一。此外,本研究发现规律光照能够显著降低内毒素血症大鼠的炎性细胞因子表达,这一结果从缓解机体炎症的角度表明了维持正常光照节律在脓毒症状态下机体加速康复中的重要意义。
本研究存在以下不足:一是持续光照刺激除影响昼夜节律调节中枢外,也能够直接影响外周组织的节律,观察到的骨骼肌消耗增加可能是中枢与外周的叠加结果;二是除持续光照外,临床上还表现为间断光照-黑暗交替模式,不同的光照节律可能对生物节律反馈环产生不同影响,需要进一步研究。
综上所述,与持续光照刺激相比,正常规律的光照可能通过维持下丘脑视交叉上核的钟基因节律表达、降低下丘脑POMC 神经肽活化来减少内毒素血症大鼠的骨骼肌消耗,对缓解脓毒症状态下的肌肉萎缩具有一定意义。