Ameliorating liver fibrosis in an animal model using the secretome released from miR-122-transfected adipose-derived stem cells
2019-11-28KeeHwanKimJaeImLeeOkHeeKimHaEunHongBongJunKwakHoJoongChoiJosephAhnTaeYunLeeSangChulLeeSayJuneKim
Kee-Hwan Kim,Jae Im Lee,Ok-Hee Kim,Ha-Eun Hong,Bong Jun Kwak,Ho Joong Choi,Joseph Ahn,Tae Yun Lee,Sang Chul Lee,Say-June Kim
Kee-Hwan Kim,Jae Im Lee,Department of Surgery,Uijeongbu St.Mary’s Hospital,College of Medicine,the Catholic University of Korea,Seoul 11765,South Korea
Kee-Hwan Kim,Ok-Hee Kim,Ha-Eun Hong,Say-June Kim,Catholic Central Laboratory of Surgery,Institute of Biomedical Industry,College of Medicine,the Catholic University of Korea,Seoul 06591,South Korea
Ok-Hee Kim,Ha-Eun Hong,Bong Jun Kwak,Ho Joong Choi,Joseph Ahn,Tae Yun Lee,Say-June Kim,Department of Surgery,Seoul St.Mary’s Hospital,College of Medicine,the Catholic University of Korea,Seoul 06591,South Korea
Sang Chul Lee,Department of Surgery,Daejeon St.Mary’s Hospital,College of Medicine,The Catholic University of Korea,Seoul 34943,South Korea
Abstract
Key words: Adipose-derived stem cells; Liver fibrosis; MicroRNAs; miR-122;Mesenchymal stem cells; Secretome
INTRODUCTION
Stem cell research is one of the promising areas of biomedical research.However,notwithstanding remarkable achievements in the field of mesenchymal stem cells(MSCs),their clinical applications are still challenging,especially due to safety concerns.To date,increasing evidence has been accumulating in support of the notion that the principal action mechanism of MSCs is secretome-mediated[1-5].Thus,to overcome the limitations of cell-based therapy,numerous researchers have focused on the exclusive use of MSC-secreted molecules rather than the cellsper se.The total set of molecules secreted or surface-shed by cells is generally referred to as secretome.The secretome includes bioactive peptides,such as cytokines,chemokines,and growth factors[1,4].These soluble factors are released from MSCs either alone or in the form of extracellular vesicles.
The therapeutic potential of secretome can be potentiated by adjusting the conditions under which MSCs are incubated.Among these conditions,the genetic modification of MSCs can offer enormous and persistent reinforcements of the MSC secretome.Literature supports that microRNAs (miRNAs) play a substantial role in the process of liver fibrosis[6-8].MicroRNAs are small non-coding RNA molecules(containing about 22 nucleotides) that alter gene expression at the posttranscriptional level,resulting in altered protein synthesis[9].Hence,miRNAs can exquisitely adjust the expression of numerous genes particularly responsible for fundamental cellular processes,such as proliferation,development,and differentiation[10].The miRNAs responsible for liver fibrosis can largely be divided into fibrotic and antifibrotic miRNAs.Of these,miR-122 is one of the representative antifibrotic miRNAs that negatively regulates collagen production in hepatic stellate cells (HSCs)[11,12].Thus,harnessing MSCs to confer miR-122 to HSCs would be a potential novel therapeutic approach for reinforcing the antifibrotic effects of MSCs.In this study,we aimed to the determine the antifibrotic effects of the secretome released from miR-122-transfected ASCs in bothin vitroandin vivomodels of liver fibrosis.
MATERIALS AND METHODS
Isolation of ASCs
Human adipose-derived stromal cells (ASCs) were obtained from lipoaspirated fat with inform consent of the volunteers.This research was approved by Institutional Review Board (IRB number 700069-201407-BR-002-01) of Hurim BioCell Co.Ltd.(Seoul,South Korea).ASCs were isolated and cultured according to previous reports[13].Lipoaspirated fat was digested by 0.1% collagenase (Sigma-Aldrich,St.Louis,MO,United States) in saline and collected after centrifugation.Cells were plated into culture flask in low-glucose Dulbecco’s Modified Eagle’s Medium(DMEM; Thermo Fisher Scientific,Hemel Hempstead,United Kingdom)supplemented with 10% FBS (Thermo Fisher Scientific),100 U/mL of penicillin(Thermo Fisher Scientific),and 0.1 mg/mL of streptomycin (Thermo Fisher Scientific).ASCs were incubated at 37 °C in humidified chamber containing 5% carbon dioxide and medium was changed every 3 d.
Transfection and attainment of secretome
ASCs were transfected with miR-122 (Exiqon,Germatown,MD) per well mixed with the Lipofectamine RNAiMAX Reagent (Thermo).After 72hr of transfection,the cells were morphologically observed by the inverted microscope.The cell numbers of the experimental groups were counted automatic cell counter (Countess®,Invitrogen,San Diego,CA,United States) using trypan blue solution.Transfected cells were processed for cell phenotyping or differentiated into three-lineage induction.
ASCs with or without miR-122 transfection were grown in a 100 mm cell dishes(Corning Glass Works,Corning,NY,United States).After reaching 70%-80%confluence,1.0 × 106ASCs were cultured in 5 mL serum-free low-glucose DMEM for 48 h.Therefore,to obtain 0.2 mL amount of secretome from 1.0 × 106ASCs,the conditioned media were concentrated 25-fold using ultra filtration units with a 3-kDa molecular weight cutoff (Amicon Ultra-PL 3; Millipore,Bedford,MA,United States).We then injected 0.1 mL amount of secretome per mouse.This means that one mouse is injected with the secretome obtained from 5 × 105ASCs.In this study,NCM refers to the secretome shed from ASCs after 48 h of incubation,and MCM refers to the secretome shed from miR-122-transfected ASCs after 48 h of incubation.
Cell phenotyping by FACS analysis
The immunophnotypes of the experimental groups were determined by flow cytometry analysis (Cytomics FC500 flow cytometer,Beckman Coulter,Fullerton,CA,United States) using FITC-conjugated CD31,CD45,and CD73 antibodies and PE-conjugated CD90 and CD105 antibodies (BD Pharmingen,San Jose,CA,United States).Isotype controls were performed with antibodies against IgG for samples.
Differentiation into adipocytes,osteocytes,and chondrocytes
Transfected cells were induced toward the three lineages for 21 d.The adipogenic,osteogenic and chondrogenic differentiation ability of MSCs was determined as previously described[14,15].Briefly,the cells were plated at a density of 1 × 104or 5 × 103cells/cm2in growth medium for 3 d,and then cultured in adipocyte and osteocyte differentiation medium (StemProTM,Gibco) for 3 wk.For chondrogenic induction,expansion medium containing 8 × 104cells was cultured for 2 h.Then,chondrogenesis differentiation medium (StemProTM,Gibco) was added and cultured for 3 wk.After differentiation,Lipid vesicles and calcium deposition were observed by oil Red O and Alizarin Red staining.For chondrogenic induction,micromass cultures were plated by seeding 5 μL droplets of 8 ×104cells into the center of 48-well plate.After incubating micromass cultures for 2 h at 37 °C,chondrogenic medium (StemPro,GIBCO) was added to 400 μL per culture wells and cultured for 3 wk.Chondrocyte induction was determined by immunohistochemical staining for collagen type I and II and proteoglycan[16].Primary antibodies were purchased from Millipore (Millipore,CA,United States) and reacted with sections.After incubation with primary antibodies,sections were incubated with PE-conjugated goat anti-rabbit immunoglobulin G (Abcam,Cambridge,MA,United Kingdom) and rabbit antimouse immunoglobulin G (Abcam).Nuclei were counterstained with DAPI (4’,6-diamidino-2-phenylindole,Invitrogen).
Human HSC culture
The LX-2 human HSCs were obtained from were kindly donated by Dr.Won-il Jeong in KAIST Biomedical research of Korea.LX-2 cells were maintained in DMEM(Thermo,Carlsbad,CA,United States).The medium was supplemented with 10% FBS(GibcoBRL,Calsbad,CA,United States),1% antibiotics (Thermo),at 37 °C.
Western blot analysis
LX-2 cells and liver specimens obtained from mice were lysed using the EzRIPA Lysis kit (ATTO Corporation; Tokyo,Japan),and quantified by Bradford reagent (Bio-RadHercules,CA,United States).Proteins were visualized by western analysis using the following primary antibodies (1:1000 dilution) at 4 °C overnight and then with HRP-conjugated secondary antibodies (1:2000 dilution) for 1 h at 25°C.From Cell Signaling Technology (Beverly,MA,United States),we obtained primary antibodies against Proliferating cell nuclear antigen (PCNA),transforming growth factor-β (TGF-β1),alpha-smooth muscle actin (α-SMA),metallopeptidase inhibitor 1 (TIMP-1),matrix metallopeptidase 2 (MMP2),collagen type- 1 alpha-1 (COL1A1),β-actin,and horseradish peroxidase (HRP)-conjugated secondary antibody.Specific immune complexes were detected using the Western Blotting Plus Chemiluminescence Reagent (Millipore,Bedford,MA,United States).
Animals and study design
Five-week male BALB/c mice (Orient Bio,Seongnam,Korea) were used in this study.Animal studies were carried out in compliance with the guidelines of the Institute for Laboratory Animal Research,Korea (IRB No:CUMC-2017-0317-04).We then compared the effects of the MCM in anin vivomodel of Thioacetamide (TAA)-induced hepatic fibrosis model.Thein vivomodel was generated by subcutaneous injection of TAA (200 mg/kg,three times a week for 8 wk) into experimental mice.Each group included 10 mice,and these were further divided into two subgroups:those for Control mice (n= 30),and those for TAA-treated mice (n= 30).Subsequently,control mice and TAA-treated mice were intravenously (using tail vein)infused with normal saline,CM,and MCM,respectively.
Serology test and ELISA
Blood samples were collected from each mouse,centrifuged for 10 min at 9500g,and serum was collected.We measured the concentrations of markers for liver injury and kidney injury,such as aspartate transaminase (AST),alanine transaminase (ALT),and creatine,using an IDEXX VetTest Chemistry Analyzer (IDEXX Laboratories,Inc.,Westbrook,ME,United States).The concentrations of mouse interleukin (IL)-6 and tumor necrosis factor (TNF)-α were measured by sandwich enzyme-linked immunosorbent assay (ELISA kits,Biolegend,San Diego,CA,United States)according to the manufacturer’s instructions.
Immunohistochemistry,Sirius red staining and masson’s trichrome staining
For immunohistochemical analysis,formalin-fixed,paraffin-embedded tissue sections were deparaffinized,rehydrated in an ethanol series and subjected to epitope retrieval using standard procedures.Antibodies against of PCNA,TIMP-1,Albumin,α-SMA,TGF-β1,MMP-2,SOD,Catalase and GPx (all from Cell Signaling Technology,MA,United States) were used for immunochemical staining.The samples were then examined under a laser-scanning microscope (Eclipse TE300; Nikon,Tokyo,Japan) to analyze the expression of PCNA,TIMP-1,Albumin,a-SMA,TGF-β1,MMP-2,SOD,Catalase and GPx.Sirius red staining and Trichrome staining were performed using the Sirius red staining kit and Masson’s trichrome staining kit according to the manufacturer’s protocol (Polysciences,Warrington,PA,United Kingdom).
Statistical analysis
All data were analyzed with SPSS 11.0 software (SPSS Inc.,Chicago,IL,United States)and SigmaPlot®ver.12.0 (Systat Software Inc.,Chicago,IL,United States).The data are presented as mean ± standard deviation (SD).Statistical comparison among groups was determined using Kruskal–Wallis test followed by Dunnett’s test as the post hoc analysis.Probability values ofP< 0.05 were regarded as statistically signif i cant.
RESULTS
Determination of differentiation potential of miR-122-transfected ASCs
We first determined whether miR-122 transfection impairs ASC functionality,especially their multilineage differentiation potential.Flow cytometric analysis showed that miR-122 transfection did not alter the expression of surface markers of ASCs (Figure 1A).Gross cell morphology was also identical regardless of miR-122 transfection (Figure 1B).In addition,transfecting miRNA did not affect multilineage differentiation potential of ASCs,including the potentials of differentiating adipocytic(Figure 1B) or osteogenic (Figure 1C) lineages,and the expression of collagens (type I and type II) and proteoglycan (Figure 1D).
In vitro experiments validating the effects of miR-122transfection into ASCs
We investigated the expression of fibrosis-related markers in miR-122-transfected ASCs.miR-122-transfected ASCs showed a decreased expression of fibrosis-related proteins (TGF β1,MMP2,α-SMA,and TIMP) compared to control ASCs or ASCs transfected with miR-122 (Figure 2A).We obtained human HSCs (LX2 cells) and treated them with a varying concentration of TAA for determiningin vitromodel of liver fibrosis.TAA elicited a concentration-dependent increase of fibrosis markers to a certain extent,and we determined that 5.0 mmol TAA is appropriate for inducing fibrosis in LX2 cells (Figure 2B and C).
Next,we obtained the secretome from the CM of ASCs as described in the method.In this study,NCM refers to the secretome shed from ASCs after 48 h of incubation,and MCM refers to the secretome shed from miR-122-transfected ASCs after 48 h of incubation.Thein vitromodel of liver fibrosis was generated by treating human HSCs cells (LX2 cells) with a hepatotoxin (TAA).We then treated the TAA-treated LX2 cells with NCM or MCM,and investigated the expression of fibrosis-related markers using western blot analysis (Figure 2D).Overall,the addition of each secretome (NCM or MCM) to TAA-treated LX2 cells significantly decreased the expression of fibrotic markers (MMP2,TGF-β1,and α-SMA) (P< 0.05).When comparing the two kinds of secretome,MCM induced the more significant reduction of fibrotic markers than did NCM (P< 0.05).
Determination of antifibrotic effects of the secretome released from miR-122-transfected ASCs in the in vivo model of liver fibrosis
We generated anin vivomodel of liver fibrosis in mouse by subcutaneous injection of TAA (200 mg/kg) three times a week for 5 wk and validated the effects of MCM in this model.The mice were divided into two groups:control (n= 30) and TAA-treated mice (n= 30),and the latter were intravenously infused normal saline (n= 10),NCM(n= 10),or MCM (n= 10) twice (200 mg/kg,three times a week for 8 wk).On the 7thd after infusion,the mice were euthanized and specimens were obtained for study.Sirius red and Masson trichrome stains were used for the estimation of fibrosis.These stains showed that,although both treatments (NCM and MCM) decreased the content of collagen,MCM significantly had the greatest effect (Figure 3A and B).In the western blot analysis of the liver specimens,MCM infusion significantly increased the expression of PCNA (a proliferation marker),and significantly decreased the expression of α-SMA,TGF-β1,and MMP1 (fibrotic markers) and increased an antifibrotic marker (TIMP-1) in the TAA-treated mice (Figure 3C).
Comparison of immunohistochemical stains
We compared the histological changes of the livers obtained from each mouse group.PCNA was used as the marker for hepatocyte proliferation; α-SMA,TGF-β1,and MMP1 for liver fibrosis; albumin for hepatic synthetic function; TIMP-1 for liver antifibrosis; and SOD,catalase,and GPx for liver antioxidant activity.Through immunohistochemical staining,the MCM group showed the highest expression of PCNA,albumin,and TIMP-1,and the lowest expression of α-SMA,TGF-β1,and MMP1 (Figure 4A and B).The MCM group also showed the highest expression of SOD,catalase,and GPx (Figure 5).
Comparison of systemic markers and liver enzymes after each treatment
We compared the expression of systemic inflammatory markers,such as IL-6 and TNF-α,in the serum of each mouse group.Secretome infusions (NCM and MCM)significantly decreased the expression of these markers,and MCM decreased their expression in a higher degree than NCM (P< 0.05) (Figure 6A).Finally,we compared the serum levels of liver enzymes (AST and ALT) in each mouse group.Secretome infusions significantly decreased the elevated levels of liver enzymes,and MCM had a higher effect than NCM (P< 0.05) (Figure 6B).
Using liquid chromatography–mass spectrometry (LC/MS),we analyzed and compared the protein contents of NCM and MCM (Figure 6C).The protein constituents and concentrations of various important proteins varied widely between NCM and MCM,validating the effects of miR-125 transfection.Specifically,MCM exhibited a significantly decreased concentration of essential intermediates of the TGF-β/Smad signaling,such as transgelin,PIN1,and profilin-1,compared to NCM.
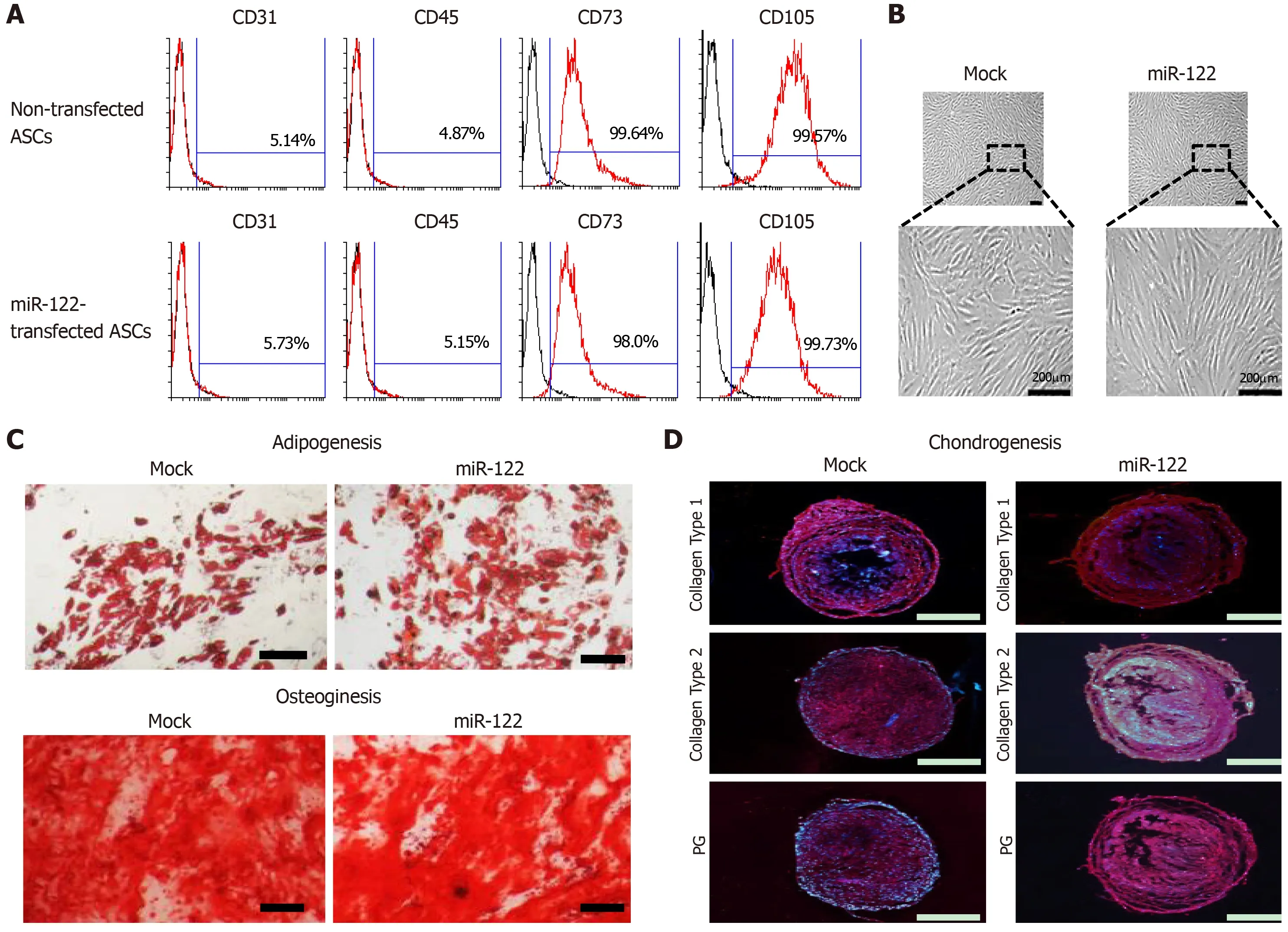
Figure 1 Assessment of multilineage differentiation potential of miR-122-transfected adipose-derived stem cells.
DISCUSSION
In this study,we have shown that the secretome released from miR-122 transfected ASCs was superior to the naïve secretome in improving liver fibrosis while minimizing inflammatory processes in mice with TAA-induced liver fibrosis.Specifically,infusion of the secretome from miR-122-transfected ASCs provided higher therapeutic potential in terms of:(A) Reducing collagen content in the liver; (B)Inhibiting proinflammatory cytokines; and (C) Reducing abnormally elevated liver enzymes than infusion of the naïve secretome.Thus,it can be postulated that miR-122 transfection into ASCs reconditions them to have higher antifibrotic properties and to release a secretome with higher antifibrotic components.In reality,our proteomic analysis of the secretome released from miR-122-transfected ASCs indicated that it had significantly lesser contents of essential intermediates of liver fibrosis compared to the naïve secretome.We could,thus,conclude that the secretome released from miR-122-transfected ASCs has higher antifibrotic and anti-inflammatory properties than the naïve secretome.
Accumulating evidence indicates that various miRNAs are essentially involved in the process of fibrosis,particularly related with the action of HSCs[17].Fibrogenic injury of the liver prompts HSCs to undergo proliferation,migrate to injured sites,and transform into myofibroblast-like cells which apparently lose their lipid droplets[18-20].Subsequently,the activated HSCs,named fibroblast-like cells,produce large amounts of extracellular matrix proteins,such as collagen I and II,fi nally leading to liver fi brosis[19,21-24].Of various cytokines,TGF-β plays essential roles in the process of liver fibrosis[25-28].
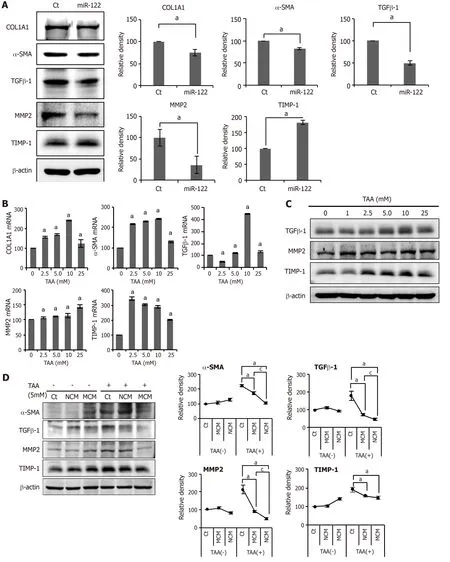
Figure 2 In vitro experiments validating the effects of miR-122 transfection into Adipose-derived stem cells.

Figure 3 Determination of antifibrotic effects of MCM in the in vivo model of liver fibrosis.

Figure 4 Immunohistochemical staining showing the effects of MCM on the expression of inflammatory and fibrotic markers in the livers.
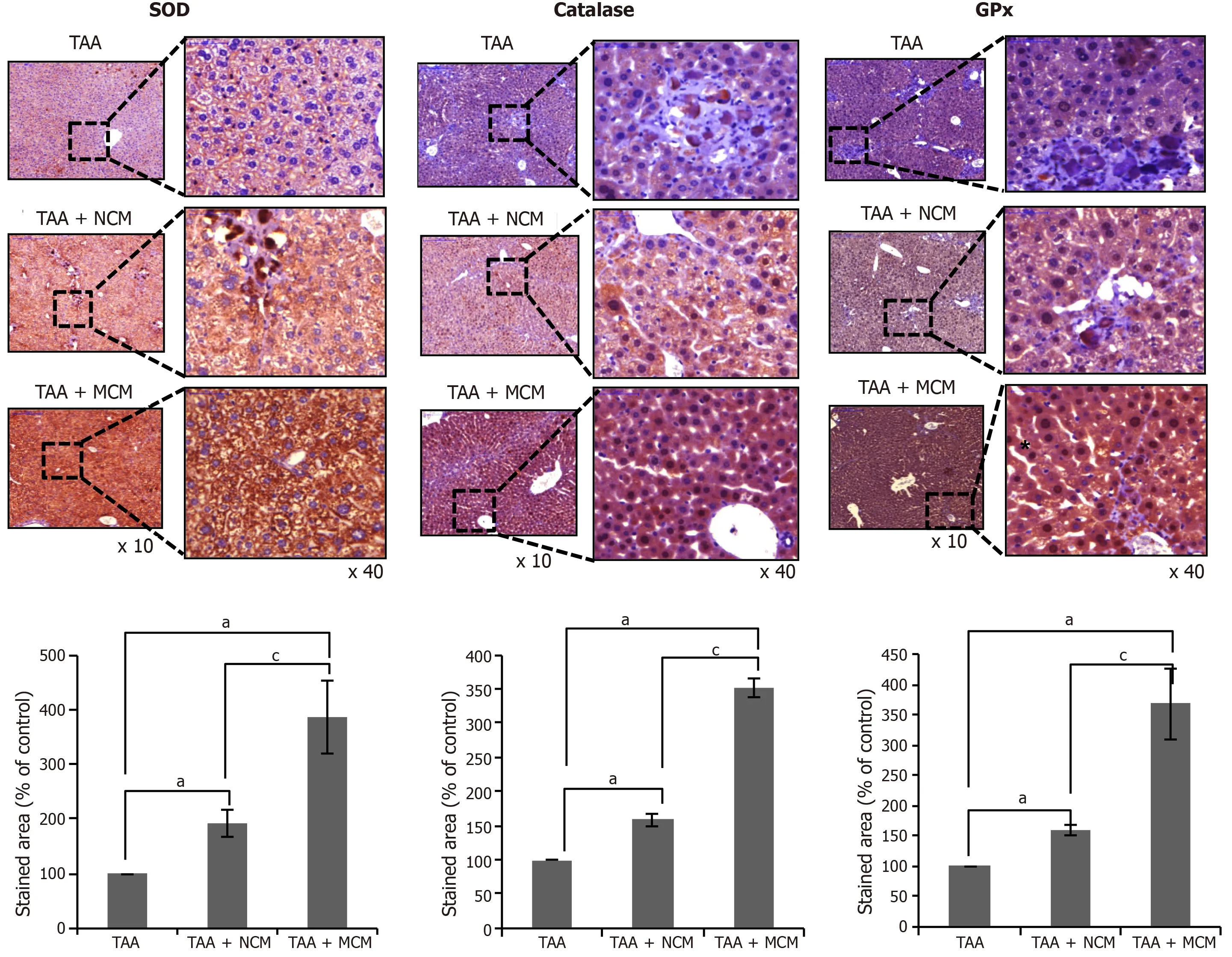
Figure 5 Effects of MCM on the expression of antioxidant enzymes in the liver.
A number of miRNAs are involved in the processes of liver fibrosis,by either promoting or preventing it.For instance,profibrotic miRNAs include miR-29b,miR-571,miR-199a,miR-200a,and miR-200b,and antifibrotic miRNAs include miR-122,miR-199,miR-200,miR-542,miR-652,and imR-181b[29-32].Specifically,miR-29b exerts its antifibrotic properties by inhibiting activation of HSCs[31].Increased serum level of miR-571 has been proposed as a potential biomarker of liver fibrosis,and serum levels of miR-542,miR-652,and imR-181b are decreased in cirrhosis.In addition,serum levels of miR-199a,miR-200a,and miR-200b were highly associated with progression of liver fibrosis in patients with chronic HCV infection[29].
miR-122 is highly expressed in liver,accounting for about 70% and 52% of total miRNAs in liver of adult mouse and human,respectively[33-35].miR-122 is essentially involved in liver development,differentiation,homeostasis,and functions.Initially,investigators revealed the crucial role of miR-122 in the regulation of cholesterol and fatty acid metabolism in the adult liver[36-38].Thereafter,anti-in fl ammatory and antifi brotic properties of miR-122 have been revealed by the generation of both germline knock-out (KO) mice and liver-speci fi c KO[39-41].Specifically,genetic deletion of miR-122 led to liver microsteatosis and in fl ammation,ultimately resulting in steatohepatitis and fi brosis[38,39].Additionally,miR-122 expression was reduced in a carbon tetrachloride-induced liver fibrosis mouse model[11].Interestingly,the restoration of miR-122 levels in miR-122 KO mice reversed the process of liver in fl ammation,by repressing two miR-122 targets,the chemokine Ccl2[39]and the profi brogenic Krüppel-like factor 6 (KLF6)[40],demonstrating potential utility of miR-122 in therapeutics.We,thus,selected the delivery of miR-122 into ASCs as a mean of reinforcing the antifibrotic properties of ASCs in this study.
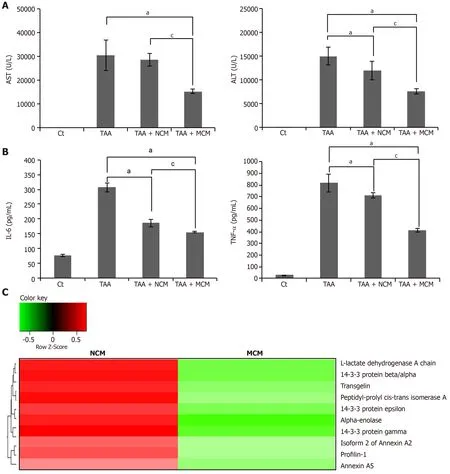
Figure 6 Determination of systemic effects of MCM and analysis of secretome components.
We have also shown that the expression of antioxidant enzymes in the liver specimens was significantly increased in the mice infused with the secretome released from miR-122 transfected ASCs compared with the mice infused with the naïve secretome.Although a variety of functional capacities of MSCs or their secretome have been reported,the protective effects against oxidative stress have rarely been reported.Kimet al[42]reported that incubation with secretomes derived from ASCs aided human dental fibroblast cells to resist free radicals,and increased antioxidant enzymes,such as SOD and glutathione peroxidase.Recently,Arslanet al[43]showed that MSC-derived exosome treatment decreased oxidative stress in the mouse model of ischemia/reperfusion.
It has been demonstrated that oxidative stress involves in both onset and progression of fibrosis arising from a variety origin,such as alcohol,viruses,iron or copper overload,or cholestasis[44].Both expression and synthesis of this inflammatory and profibrogenic cytokines are mainly modulated through redox-sensitive reactions[45,46].Further,redox-sensitive reactions also involve in other essential processes of liver fibrosis,such as activation of HSCs and expression of metalloproteinases and of their specific inhibitors[47-49].We thus think that reduction of oxidative stress could be another way of antifibrotic mechanisms exerted by the secretome released from miR-122 transfected ASCs.
Here,we have focused on the effects of secretome and not those of the stem cells,on liver fibrosis.The term secretome was first mentioned by Blacket al[50]to refer to all the factors secreted by a cell,along with the secretory pathway constituents.The main constituents of a secretome include secretory proteins and extracellular vesicles.The secreted proteins in humans account for 13%-20% of the entire proteome and include growth factors,cytokines,chemokines,adhesion molecules,proteases,and shed receptors[51].Extracellular vesicles are typically 30-2000 nm in diameter and can be subdivided into exosomes,microvesicles,and apoptotic bodies,according to their size.Extracellular vesicles usually contain and,thus,carry non-protein components,such as lipids,DNAs,micro-RNAs,and mRNAs.In this study,we focused on the effects of the whole secretome,not its individual constituents,such as exosomes.Exosomes,for example,can be obtained by protracted,complex,and expansive processes[52].We expect that our results will help eliminate the laborious and expensive process of obtaining exosomes.
The concept of using miRNAs for enhancing the therapeutic potential of the secretome released from stem cells is quite different from how they have been used before.Previously used methods for potentiating secretome,which include physical and chemical stimulation methods,such as hypoxic preconditioning[53,54]or the use of lipopolysaccharides[55],can be categorized as nonspecific stimulation.By contrast,the concept of using miRNAs can be categorized as liver-specific stimulation.In the future,the clinical application of secretome is expected to be tailored according to the needs of patients,combining nonspecific and specific stimulations.
In conclusion,we have shown that the secretome released from miR-122-transfected ASCs was superior to the naïve secretome in improving liver fibrosis,while minimizing inflammatory processes,in mice with TAA-induced liver fibrosis.Hence,it can be postulated that miR-122 transfection into ASCs reconditioned them to have higher antifibrotic properties and to release a secretome with higher antifibrotic components.We could,thus,conclude that the secretome released from miR-122 transfected ASCs has higher antifibrotic and anti-inflammatory properties than the naïve secretome.Because miR-122 transfection into ASCs provides a specific way of potentiating the antifibrotic properties of the ASC secretome,it could be considered as an enhanced method of reinforcing secretome effectiveness.
ARTICLE HIGHLIGHTS
Research background
The therapeutic potential of mesenchymal stem cells (MSCs) is known to be mediated mainly by the secretome that refers to the total collection of secretory materials from MSCs.Basically,naïve secretome has anti-inflammatory,immunomodulatory,and tissue reparative properties.To increase the amount or to reinforce the potential of naïve secretome,researchers have attempted to adjust physico-chemical environment of MSCs or genetically manipulate MSCs.The former has the advantage of being simple but lacking persistence,while the latter has a strong persistence but has the disadvantage of a safety concern in the clinical application.
Research motivation
We have been considering genetic modification as a way of persistently potentiating the therapeutic potential of naïve secretome.In addition,contrasted by the use of genetically modified MSCs,we thought that the use of the secretome could significantly lower the safety concern.We also noted miRNAs as the materials to be used for genetic manipulation,because miRNA is critically involved in the process of liver fibrosis.
Research objectives
Our aim was to determine the antifibrotic potential of the secretome released from miR-122-transfected adipose-derived stromal cells (ASCs) in the model of liver fibrosis.
Research methods
Secretory materials released from ASCs that had been transfected with antifibrotic miR-122 were collected and termed as miR122-secretome.Thein vitromodel of liver fibrosis was generated by treating human hepatic stellate cells (LX2 cells) with a hepatotoxin (thioacetamide; TAA),and thein vivomodel of liver fibrosis was generated by subcutaneous injection of TAA (200 mg/kg,three times a week for 8 wk) into five-week male BALB/c mice.For determiningin vivoeffects of miR122-secretome,each secretome (miR122-secrectome and naïve secretome) was intravenously administered to the mice with liver fibrosis,respectively.The degree of liver fibrosis and other alternations in cells or tissues were determined using by molecular and histological investigations,including cell viability assay,western blotting,immunohistochemistry,serology tests,and sandwich enzyme-linked immunosorbent assays.
Research results
The addition of miR-122-secretome to fibrosis-induced LX2 cells significantly decreased the expression of fibrotic markers (MMP2,TGF-β1,TIMP-1,and α-SMA) and increased the expression of an antifibrotic marker (TIMP-1).The western blot analysis showed that miR122-secretome infusion significantly increased the expression of PCNA (a proliferation marker),significantly decreased the expression of α-SMA,TGF-β1,and MMP1 (fibrotic markers),and increased an antifibrotic marker (TIMP-1) in the livers of TAA-treated mice.In addition,miR122-secretome infusion significantly reduced collagen content in the livers,inhibited serum levels of proinflammatory cytokines,such as IL-6 and TNF-α,as well as serum levels of liver enzymes than infusion of the naïve secretome.Finally,our analysis of the components of miR-122-secretome showed that miR-122-secretome exhibited a significantly decreased concentration of essential intermediates of the TGF-β/Smad signaling,such as transgelin,PIN1,and profilin-1,compared to NCM.
Research conclusions
miR-122-secretome was found to be superior to the naïve secretome in improving liver fibrosis while minimizing inflammatory processes in mice with TAA-induced liver fibrosis.Our proteomic analysis of the miR-122-secretome also validated that miR-122-secretome had significantly lesser contents of essential intermediates of liver fibrosis.Therefore,transfecting miR-122 into ASCs is worth considering as a way of reinforcing antifibrotic properties of the secretome from ASCs.
ACKNOWLEDGEMENTS
We thank Byung-Rok Do and Ji-Hyang Kim in Hurim BioCell Company for providing human ASCs and technical assistance.We would like to thank the Francis Sahngun Nahm (a professional statistician) for his devoted assistance of statistical analysis.We would like to thank Hye-Jung Kim for photoshop works that improved the figure quality.We also would like to thank Ji-Hye Park for data processing and statistical analysis.We would like to thank Drug and Disease Target Team (Ochang),Korea Basic Science Institute.
杂志排行

World Journal of Stem Cells的其它文章
- Unexpected encounter of the parasitic kind
- Colon cancer stemness as a reversible epigenetic state: Implications for anticancer therapies
- CRISPR/Cas system: An emerging technology in stem cell research
- Cytokine interplay among the diseased retina,inflammatory cells and mesenchymal stem cells - a clue to stem cell-based therapy
- Developments in cell culture systems for human pluripotent stem cells
- Monitoring maturation of neural stem cell grafts within a host microenvironment