共轭亚油酸对猪肉品质的影响及调控机制研究进展
2019-11-27陈文涛徐子叶单体中
陈文涛,徐子叶,单体中
(浙江大学动物分子营养学教育部重点实验室,浙江大学饲料科学研究所,浙江杭州 310058)
我国是世界上最大的猪肉生产国和消费国。在过去30 余年,通过大量的良种引进、营养水平和环境条件改善等措施,猪的生长速度和饲料转化效率得到大幅度提高,猪的产肉性能也得到了很大提升,猪肉产量已基本能满足人们的需要。然而,随着国民经济的发展和人民生活水平的提高,消费者越来越关注肉品质、口感、风味和营养等,猪肉品质问题已成为养猪业发展亟需解决的关键问题之一。因此,如何生产安全优质的猪肉产品是目前畜牧工作者研究的热点。
猪肉品质的评价指标主要包括肉色、嫩度、风味、大理石纹、肌内脂肪含量和脂肪酸组成等,其影响因素主要包括品种、营养、环境等。其中,营养调控是目前改善和提高猪肉品质的有效措施之一。共轭亚油酸(Conjugated Linoleic Acid,CLA)作为一类必需脂肪酸,具有多种生物学功能,在调控糖脂代谢、免疫、癌症发生和肌肉及骨骼发育等方面均发挥重要作用[1]。在养猪生产中,CLA 可以降低动物机体体脂沉积、提高肌内脂肪含量、改变脂肪酸组成以及影响肌纤维类型等。因此,本文在阐述CLA 的结构和功能的基础上,重点论述了CLA 对猪肉品质的影响及可能的调控机制,为通过营养调控策略改善猪肉品质提供一定的理论依据。
1 CLA 的结构与功能
1.1 CLA 的结构与来源 CLA 是一类含有18 个碳原子和2 个双键的脂肪酸的总称。CLA 主要有2 个来源:一个是反刍动物瘤胃细菌发酵,是氢化亚油酸或亚麻酸为硬脂酸的中间产物;另一个是通过δ-9-去饱和酶对异柠檬酸的去饱和作用,在反刍动物乳腺中内源性形成[2]。在这些异构体中,反式-10(t10)、顺式-12(c12)和顺式-9(t9)、反式-11(c11)CLA 因其广泛的生物效应而成为研究重点。这2 种异构体在结构、来源和功能方面均有显著差异。比如,天然存在的异构体80%以上均是c9-t11-CLA,t10-c12-CLA 主要来自于化学合成,天然存在含量很低。除了具有抗癌和抗动脉粥样硬化的生物活性,这2 种异构体都有一些独特功能。c9-t11-CLA 对预防和抑制癌症发生和肿瘤生长的效果更显著,t10-c12-CLA 则可降低体脂、提高瘦肉率和调节脂肪酸组成[3]。
1.2 CLA 的代谢 CLA 在生物系统内的代谢过程与脂肪酸一致。饲粮中的CLA 在脂肪酶作用下分解,经过肠道的消化吸收和血液循环运输到肝脏等组织,在体内进行利用。补充的CLA 主要沉积在甘油三酯和磷脂中,CLA 的代谢产物也能在动物组织中检测到[4]。然而,由于c9-t11-CLA 和t10-c12-CLA 结构不同,在代谢水平上也存dkkdddd12 异构体抵制C20:3 和C20:4 形成,生成C18:3[5],但CLA 的几何结构和双键位置不影响其在小肠内吸收[6]。此外,CLA 不会扰乱所有非共轭不饱和脂肪酸的代谢,仅影响其代谢产物沉积到中性脂质和磷脂中的比例[7]。中性脂质比例的差异间接影响不同组织中脂肪酸的组成模式和脂肪含量,而磷脂是生物体内代谢的重要调控成分。可见,CLA 异构体间的差异性主要来自于不同组织和细胞沉积异构体的能力。
1.3 CLA 的生理功能
1.3.1 调节糖脂代谢 糖脂代谢是细胞的基本生命活动之一,CLA 与脂肪合成与分解、糖原的利用和分配等糖脂代谢密切相关。t10-c12-CLA 处理可以降低内脏、腹股沟等脂肪沉积,增加血浆葡萄糖浓度,提高白色脂肪的分解代谢活性[8],提示CLA 可通过促进脂质分解降低脂肪沉积。另外,CLA 可以作用于骨骼肌和肝脏等代谢器官,通过激活过氧化物酶体增殖物激活受体γ(Peroxisome Proliferators-Activated Receptor Gamma,PPARγ)途径上调脂联素的表达,改善胰岛素敏感性,降低血糖浓度,调控糖代谢稳态[9]。
1.3.2 抗氧化作用 CLA 能清除体内自由基,增强体内抗氧化酶系统活性。CLA 处理后的大鼠血液和肝脏中过氧化氢酶(Catalase,CAT)、超氧化物歧化酶(Superoxide Dismutase,SOD)和谷胱甘肽转移酶(Glutathione S-transferase,GST)酶活性大幅度增加[10]。CLA 的抗氧化作用在家禽上也得到证实。吴国玲[11]研究发现,补充CLA 可显著影响鸡血清中总超氧化物歧化酶(Total Superoxide Disumutase,T-SOD)和谷胱甘肽过氧化物酶(Glutathione Peroxidase,GSH-Px)水平;随着CLA 添加量增加,T-SOD 和GSH-Px 呈先升高后降低的趋势,并且CLA 的抗氧化性能优于维生素A。
1.3.3 抗炎反应 CLA 在多种炎症动物模型中发挥有益作用。CLA 能降低人肺癌细胞中肿瘤坏死因子-α(Tumor Necrosis Factor-Alpha,TNF-α)、白 细 胞介素-1β(Interleukin 1 Beta,IL-1β)和前列腺素E2(Prostaglandin E2,PGE2)的产生,但对白细胞介素6(Interleukin 6,IL-6)的产生没有显著影响[12]。CLA 可减少牛乳腺上皮细胞中大肠杆菌引起的炎症反应,这种作用是通过TLR4-NF-κB 和PPARγ 途径参与介导的[13]。另外,膳食补充CLA 缓解了葡聚糖硫酸钠诱导的结肠炎引起的体重减轻和直肠出血,这种有益效应与PPARγ、过氧化物酶体增殖物激活受体γ 共激活因子1α(Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha,PGC1-α)和TNF-α表 达变化有关[14]。猪饲料中添加CLA 减轻了由脂多糖(Lipopolysaccharides,LPS)刺激诱导的生长抑制和IL-6 和TNF-αmRNA 表达的升高[15]。
1.3.4 抗癌作用 CLA 对前列腺癌细胞、乳腺癌细胞、结肠癌细胞和胃癌细胞等多种肿瘤细胞均有显著抑制作用[16],抗癌效果与CLA 的剂量有关。c9-t11-CLA 和t10-c12-CLA均能抑制体内血管生成[17],通过降低血清中血管内皮生长因子(Endothelial Growth Factor,VEGF)和碱性成纤维细胞生长因子(Fibroblast Growth Factor,FGF)水平以及阻断胎肝激酶1(Fetal Liver Kinase-1,FLK-1)受体发挥其抗癌作用,从而抑制对肿瘤生长和存活至关重要的血管生长[18]。诱导细胞凋亡也是抗癌的重要途径,CLA 可由蛋白激酶B/糖原合成酶激酶3β(Protein Kinase B,AKT/Glycogen Synthase Kinase3β,GSK3β)和腺苷酸激活蛋白激酶(Adenosine 5´-Monophosphate(AMP)-Activated Protein Kinase,AMPK)途径诱导细胞凋亡,发挥抗肿瘤的生物活性[19-20]。CLA 抗癌作用涉及了多种细胞活动,抑制细胞增殖、诱导细胞凋亡和调节血管生成是其发挥抗癌作用的关键途径。
1.3.5 调控骨骼发育 骨骼与能量代谢以及葡萄糖和脂肪调控密切相关。饲喂高脂肪饮食会导致骨骼形成受损,饮食中添加CLA 会对小鼠和细胞中的骨代谢产生积极影响[21]。CLA 能够调节参与骨骼生成和能量代谢的基因的表达,进而对骨代谢发挥有益作用[22]。CLA 处理后,前成骨细胞中碱性磷酸酶(Alkaline Phosphatase,ALP)、runt 相关转录因子2(Runt-Related Transcription Factor 2,RUNX2)和骨钙素(Osteocalcin,OCN)表达上调,说明CLA可以通过AKT/哺乳动物雷帕霉素靶蛋白(Mammalian Target Of Rapamycin Complex 1,mTORC1)信号途径刺激成骨细胞的分化[23]。Rahman 等[24]研究表明,CLA 通过显著抑制核因子受体活化蛋白配体(Receptor Activator Of Nuclear Factor Kappa-B Ligand,RANKL)诱导的核因子-κB(Nuclear Factor-Kappa B,NF-κB)活化途径来抑制破骨细胞生成。
2 CLA 对猪肉品质的影响
2.1 CLA 对猪肉感官品质的影响 肉的感官品质是肉质评价的重要参考,包括肉色、系水力、嫩度、风味和pH。CLA 经过体内的消化、吸收、代谢和沉积过程差异性地影响肉的生理生化指标,进而引起猪肉感官品质的变化。
许多研究证实了CLA 在提高猪肉感官品质方面的有益效果。刘倩倩等[25]发现,CLA 能提高猪肉的肉色评分和大理石花纹评分,但对猪肉的嫩度、系水力、风味及pH 等无显著影响。王琪等[26]报道,饲粮中持续添加1.5% CLA 能显著提高育肥猪眼肌面积、肌内脂肪、肉色评分和大理石纹评分,极显著提高屠宰重、肌肉pH。Barnes 等[27]研究发现日粮中添加CLA 有增加大理石纹的趋势,CLA 对大理石花纹的影响可能与增加肌肉内脂肪细胞大小有关。Joo 等[28]也发现,饲喂 CLA 日粮可增加猪肌内脂肪含量和系水力,同时抑制肌肉的脂质氧化,提高肉色稳定性。但CLA 的作用效果受CLA 的添加剂量和时间及猪品种、性别和日龄等多种因素影响。
2.2 CLA 对猪肉中脂肪酸组成的影响 脂肪酸作为一种能源物质,不仅是动物细胞调节和新陈代谢不可缺少的成分,同时决定了脂肪的物理和质地特性以及肌肉的氧化稳定性,对肉类的营养价值起着关键作用。CLA 能增加组织中脂肪酸的饱和度,并且提高多不饱和脂肪酸(Polyunsaturated Fatty Acids,PUFA)与单不饱和脂肪酸(Monounsaturated Fatty Acids,MUFA)的比例[29]。Cordero 等[30]报道,日粮中补充CLA 不仅线性增加猪背最长肌中肌内脂肪含量,同时增加肌肉和脂肪组织中饱和脂肪酸(Saturated Fatty Acid,SFA)的水平,降低MUFA 的水平。Tous 等[31]研究发现,饲粮中添加CLA 提高母猪背最长肌、半膜肌、肝脏及皮下脂肪的SFA 比例,降低皮下脂肪和背最长肌MUFA 及肝脏、半膜肌和皮下脂肪的PUFA 水平,表明CLA 调控脂肪酸组成的效果具有高度组织特异性。
此外,补充CLA 不仅调控机体组织中脂肪酸组成,而且可以改变乳脂的合成和脂肪酸的比例。Peng等[32]研究发现,补充CLA 影响母乳脂肪酸组成,膳食CLA 增加了总SFA 的浓度,同时降低了初乳中的总MUFA,但不影响总MUFA 浓度。Krogh 等[33]试验评估了膳食CLA 对初乳的产量和组成、初产期和母猪产奶量的影响,结果表明CLA 改善了初乳中乳脂率和脂肪酸组成,并刺激产奶量的提高,但不影响初乳中蛋白质、干物质和乳糖的含量。可见,CLA 在母体的沉积效果会通过乳汁传递给后代。
3 CLA 调控猪肉品质的机制
影响猪肉品质的直接关键因素是脂肪(肌内脂肪)沉积和肌肉发育(肌纤维类型);而糖脂代谢是最主要的生命活动之一,与脂肪沉积、肌肉发育和肉品质形成密切相关。因此,CLA 对猪肉品质的调控机制可归纳为3 条途径:①CLA 对脂肪沉积的调控机制;②CLA对肌肉发育(肌纤维类型)的调控机制;③CLA 对糖脂代谢的调控机制(图1)。无论哪种途径,CLA 在体内的生理反应都离不开动物机体中相关基因及相关信号通路的参与。
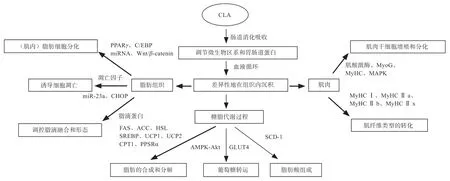
图1 CLA 调控猪肉品质形成的作用机制
3.1 CLA 对脂肪沉积的调控机制
3.1.1 CLA 抑制脂肪细胞的分化 脂肪前体细胞经过增殖和分化过程才能转化为成熟的脂肪细胞,CLA 通过调控脂肪分化关键基因的表达影响脂肪沉积量。PPARγ和CCAAT/ 增强子结合蛋白(CCAAT/enhance Binding Proteins,C/EBPs)是调控脂肪细胞分化的关键基因。Kang 等[34]报道,t10-c12 CLA 显著抑制脂肪组织中PPARγ的表达,体脂增加明显减少。CLA 处理导致3T3-L1 前脂肪细胞中C/EBPα和PPARγ的表达下调[35],说明CLA 会抑制前体脂肪细胞分化成脂肪细胞。调控脂肪分化的信号通路的活性,可调控下游靶基因表达,也能调控脂肪细胞的分化进程。Wnt/β-catenin 是一条抑制脂肪生成的重要信号通路。Yeganeh 等[36]研究发现,t10-c12-CLA 通过影响3T3-L1 脂肪细胞中的Wnt/β-catenin 途径来抑制脂肪细胞分化,增强PPARγ在无活性复合物中的螯合,从而防止脂肪生成。
CLA 调控分化的途径是复杂多样的,除了通过调控脂肪细胞分化关键基因的表达水平外,CLA 也影响相关miRNA 表达。Wang 等[37]采用高通量测序方法研究CLA 对脂肪组织miRNA 的影响,筛选并鉴定出参与Wnt 信号传导途径的14 种miRNA,并通过影响脂肪细胞分化途径和改变胞内信号传导途径来调控体脂沉积。这与Qi 等[38]研究结果相同,即CLA 通过调节脂肪组织中的miRNA 表达来降低体脂沉积。上述研究结果表明,CLA 可通过调控miRNA,调控下游靶基因的表达,进而影响脂类代谢和肉品质形成。
另外,CLA 可以差异性调控猪皮下脂肪前体细胞和肌内脂肪前体细胞的分化进程。研究发现,CLA 可降低皮下脂肪含量,促进肌内脂肪沉积,改善胴体品质,其机制是 CLA 通过降低皮下脂肪细胞特异性基因的表达以及脂肪前体细胞数量,提高肌内脂肪组织特异性基因的表达促进肌内脂肪沉积[39]。
3.1.2 CLA 诱导脂肪细胞凋亡 激活脂肪细胞凋亡途径,导致细胞增殖和凋亡过程的失衡是CLA 调控脂肪沉积的又一途径。Corino 等[40]报道,CLA 处理后猪的前体脂肪细胞增殖率降低,而脂肪细胞凋亡数量增多。然而,CLA 诱导体脂肪减少和脂肪细胞凋亡的作用具有异构体特异性。Zhai 等[41]在体外用高剂量的CLA 长期处理培养的3T3-L1 细胞,发现t10-c12-CLA 能刺激脂肪细胞凋亡,但c9-t11-CLA 不会促进脂肪细胞凋亡。Qi 等[42]研究发现,CLA 处理显著下调miR-23a 的表达,进而通过凋亡蛋白酶激活因子1(Apoptotic Protease Activating Factor-1,APAF1)诱导脂肪细胞的凋亡。t10-c12-CLA 处理后的小鼠会促进TNF-α和IL-6基因表达[43],而TNF-α 在诱导细胞凋亡过程中发挥重要功能。Ou 等[44]报道,CLA 处理增加C/EBP 同源蛋白(C/EBP Homologous Protein,CHOP)表达和内质网应激反应,导致细胞凋亡。Qi 等[45]在猪上的试验证实,CLA 会降低抗凋亡因子Bcl-2 的表达,增加促凋亡因子Bax 和P53 的表达,进而促进脂肪细胞凋亡来减少猪体脂沉积。上述结果表明,CLA 可以通过不同机制诱导脂肪细胞凋亡进而影响脂质代谢。
3.1.3 CLA 调控脂滴形成 脂滴是存储中性脂质的胞质细胞器,对脂肪沉积及能量代谢至关重要,CLA 可通过调控脂滴的形态和数量来调节甘油三酯的储存。t10-c12-CLA 可以减少小鼠白色脂肪组织和体外培养的脂肪细胞中的脂滴;进一步研究发现,t10-c12-CLA 可增加脂解和改变脂肪细胞脂滴形态,而不是c9-t11-CLA[46]。另外,t10-c12-CLA 会改变猪卵母细胞成熟过程中细胞质脂滴的分布和形态,从而以时间依赖性方式降低其脂质含量[47]。
CLA 调控脂滴形成的过程实际上是通过脂滴蛋白发挥作用。t10-c12-CLA 介导的脂肪细胞甘油三酯含量的减少与脂滴相关蛋白的差异定位和表达相关,该过程通过激活mTOR/p70S6K/S6 信号传导实现对脂肪分化相关蛋白(Adipocyte Differentiation Related Protein,ADRP)的翻译控制和对Perilipin A 的转录控制[48]。Yeganeh 等[49]报道,t10-c12-CLA 激活PKCα 途径调控脂滴的数量和脂滴蛋白的磷酸化水平,进而调控脂肪沉积。另外,Stringer 等[50]利用Zucker 大鼠(胰岛素抵抗和代谢综合征的模型)评估CLA 对肝脏脂肪变性的作用,结果表明c9-t11-CLA 处理的肝细胞中具有更少的细胞质脂滴,但是这些脂滴的尺寸更大;而t10-c12-CLA 处理的肝细胞具有更多数量的脂质小滴,且脂滴的变化与较低的Perilipin 2 水平相关。
3.2 CLA 对肌肉发育(肌纤维类型)的调控机制 肌纤维是肌肉的基本组成成分,肌肉的生长发育与肌纤维的长度、直径和类型密切相关。肌纤维的数量在胚胎期已经确定,出生后不再改变,肌纤维直径和长度依赖于肌卫星细胞的发育,肌纤维类型的转化则伴随着整个机体发育时期。CLA 可通过改变与肌肉生长发育相关的基因表达来调节肌纤维组成和类型。体外研究表明,c9-t11-CLA 处理可提高C2C12 细胞的增殖和分化,而t10-c12-CLA 显著抑制细胞增殖和分化进程[51],表明不同CLA 异构体对肌肉生长存在差异性调控。此外,用t10-c12-CLA 孵育C2C12 细胞,降低了肌肉特异性基因肌酸激酶、肌细胞生成素、肌球蛋白重链的表达,从而抑制了肌管的形成[52]。
肌纤维的组成和比例与肉品质有密切关系,肌球蛋白重链I(Myosin Heavy Chain I,MyHC I)和MyHC Ⅱa 比例越高,猪肉的pH、肉色、嫩度和风味等与肉品质相关的指标就越高。体内外试验均证明了CLA 在促进肌纤维转化方面的功能。在日粮中添加1.5%的CLA,生长肥育猪的MyHC I 和MyHC Ⅱa mRNA 表达量显著高于对照组,而MyHC Ⅱb 和MyHC Ⅱx mRNA 表达量显著低于对照组[53]。Huang 等[54]研究也发现,CLA显著影响肌纤维型基因的表达,显著提高MyHC I 和MyHC Ⅱa 表达,下调MyHC Ⅱx 表达,但对MyHCⅡb mRNA 表达影响不显著。另外,体外培养的猪骨骼肌卫星细胞向肌纤维转化时,添加不同水平CLA 均能提高MyHC I 和MyHC Ⅱa 型肌纤维比例,降低MyHC Ⅱb 和MyHC Ⅱx 型肌纤维比例[55]。重要的是,膳食CLA 还可以缓解生长肥育猪由于年龄变化引起的MyHC I mRNA 丰度下降的趋势,增加氧化型纤维的比例[56]。上述研究揭示,CLA 可以通过调控肌肉发育和肌纤维类型进而影响产肉性能和肉品质的形成。
3.3 CLA 对糖脂代谢的调控机制
3.3.1 CLA 影响脂类代谢关键酶的活性 脂类代谢是一个极其复杂的生理过程,包括脂肪酸的摄取、转运、延伸及氧化,脂肪的合成、储存与分解等多个生物学过程。补充CLA 可通过调控与脂类代谢关键酶的转录来影响脂肪沉积。膳食CLA 降低了肌肉和肝脏中与脂肪合成相关酶的活性,进而改变组织中脂肪酸组成和脂肪含量[57]。José 等[58]报道,t10-c12-CLA 降低了脂肪酸合成酶(Fatty Acid Synthase,FAS)活性,但对猪脂肪组织的脂质分解没有影响。而Ostrowska 等[59]研究发现,CLA 显著增加血浆非酯化脂肪酸和甘油三酯的浓度,表明脂肪动员和脂质分解增加,推测可能是脂蛋白脂酶活性变化引起的。参与葡萄糖和脂肪酸生物合成的基因转录水平显著降低,包括乙酰辅酶A 羧化酶-α和-β(Acetyl-coa Carboxylase,ACC)、激素敏感脂肪酶(Hormone-Sensitive Lipase,HSL)、脂蛋白脂酶(Lipoprotein Lipase,LPL)和固醇调节元件结合蛋白(Sterol Regulatory Element Binding Protein 1,SREBP-1),而解偶联蛋白(Uncoupling Protein 1,UCP1)、肉碱棕榈酰转移酶(Carnitine Palmitoyl Transferase 1,CPT1)mRNA水平增加[60]。这表明补充CLA 会通过增强脂肪酸氧化来增加储存的甘油三酯的消耗,减少脂肪合成。Wang等[61]用t10-c12-CLA 处理培养的脂肪细胞,线粒体呼吸和脂肪酸氧化增加。Choi 等[62]证实,补充c9-t11-CLA可增加肝脏脂联素受体1 和2 的敏感性,导致PPARα配体活性和脂肪酸氧化增加。这说明CLA 在体内一方面促进组织脂肪酸氧化和分解,加速能量利用;另一方面减少脂肪的合成,从而使体脂沉积减少。
CLA 异构体和其代谢物参与不同组织的磷脂和中性脂质部分,显著改变了脂肪酸谱系。House 等[63]报道,脂肪酸间比例和组成的变化归因于硬脂酰辅酶A 去饱和 酶-1(Stearoyl-Coenzyme A Desaturase 1,SCD-1)活性的降低,其结果进一步说明CLA 的作用效果和路径与作用时间有关。Jiang 等[64]报道得到了类似的结论,补充CLA 降低背膘和背最长肌中的△9-去饱和酶活性。研究CLA 对新生仔猪大脑和肝脏组织中n-6 长链多不饱和脂肪酸代谢过程中发现,膳食CLA 对脂肪酸β-氧化没有影响,但通过抑制伸长和去饱和作用可显著降低n-6长链多不饱和脂肪酸生物合成[65]。上述研究表明,CLA通过抑制脂肪酸去饱和酶的活性来调控脂肪酸组成。
3.3.2 CLA 调控葡萄糖的转运 骨骼肌通常占总体重的近40%,在糖脂代谢和整体能量稳态中发挥重要的调节作用。在基础条件下,CLA 补充会促进小鼠肌肉中葡萄糖的氧化和利用[66],这增加了葡萄糖摄取和肌肉中葡萄糖的转运。可能是由于胰岛素受体和葡糖糖转运蛋白数量的增加,进而允许更高剂量的葡萄糖跨越肌细胞膜用作能量来源。在骨骼肌细胞中,CLA 通过磷脂酰肌醇3(Phosphatidylinositide 3-Kinase,PI3)激酶和AMPK 途径,刺激葡萄糖转运蛋白-4(Glucose Transport Proteins 4,GLUT4)易位至质膜促进葡萄糖摄取[67]。也有研究报道,t10-c12-CLA 通过Ca2+-CaMKII-AMPK-Akt途径以刺激骨骼肌葡萄糖转运[68]。高水平的葡萄糖进入肌肉细胞中,肌内脂肪供能减少,保留了肌内脂肪酸储备,肉品质得到改善。
CLA 在调控代谢综合征动物模型中葡萄糖的利用也承担了重要作用。t10-c12-CLA 处理可改善肥胖大鼠中骨骼肌葡萄糖耐受性和葡萄糖转运活性的能力,这种变化与氧化应激和肌肉脂质水平的降低有关[69]。CLA不仅对母体有效果,而且对具有代谢综合征母体的后代同样发挥作用。CLA 补充缓解了血浆中的高胰岛素和高葡萄糖浓度,进而改善了母体高脂饮食诱导的大鼠后代代谢功能障碍[70]。
3.3.3 CLA 影响肠道菌群 胃肠道整合了食物、微生物群和宿主代谢之间的相互作用,将外源性的营养素经过肠道消化吸收,构成了体内各器官生长发育的基础,对机体产生有益代谢反应。已经鉴定出肠道内发酵可产生CLA 的菌群,重要的是,提升产CLA 有益菌属的丰度一方面影响了盲肠和肝脏组织中CLA 的水平,另一方面可以降低动物模型中结肠癌的发病率和减轻炎症及脂类代谢[71-72]。而CLA 异构体经过肠道的消化也会影响微生物区系组成,进而调节体内脂质代谢过程。在高脂饮食下补充CLA 对小鼠胃肠道的特定部位有显著影响,增加多种胃蛋白表达和促进肠道有益微生物群的定植[73]。补充t10-c12-CLA 后小鼠肠道微生物组成发生改变,并且缓解了肥胖小鼠的体重增加,推测肠道微生物的变化与体内能量代谢水平增加有关,进而引起体重减少[74]。研究发现,t10-c12-CLA 处理可显著改变与脂质代谢和诱导肝脏脂肪变性肠道微生物组成,厚壁菌门比例降低,拟杆菌比例升高,但没发现c9-t11-CLA 具有同样的效果[75]。这些结果表明,饮食t10-c12-CLA 对小鼠脂质代谢的调控,可能部分由肠道微生物群组成和功能的改变介导。
4 小结与展望
综上所述,CLA 可以通过调控脂肪细胞分化、糖脂代谢和肌肉发育(肌纤维类型)来影响肌内脂肪沉积、肌肉发育和肌纤维类型,进而影响脂肪酸组成、肉色、系水力、风味、嫩度和pH 等肉的感官品质和营养品质。因此,CLA 作为一种绿色安全的饲料添加剂,在养猪业和优质安全猪肉生产中具有广泛的应用前景。然而,目前CLA 在生产应用上仍然存在一些问题:CLA 的作用效果不稳定,尤其是CLA 在不同品种、日龄、性别畜禽中应用的效果差异显著;CLA 在肠道内消化吸收后,如何调控体内器官和组织发育的过程和途径不明确;CLA 吸收后在不同组织内的利用和分配也不清晰,无法实现精准调控;CLA 的制备工艺和成本较高,无法大规模推广应用。另外,CLA 在体内的生理功能和调控机制错综复杂,是多种细胞活动和代谢途径相互作用的结果。因此,关于CLA 对肉品质形成的作用调控机制仍需要深入研究,进而为CLA 的广泛应用和优质猪肉生产调控提供理论和实践依据。
