牡丹籽蛋白酶解产物对禽多杀性巴氏杆菌ptfA基因DNA疫苗免疫效果的影响
2019-11-22杜珍奇牛明福秦翠丽任国艳马丽苹伍家发
杜珍奇,宫 强,牛明福,秦翠丽,任国艳,马丽苹,伍家发,张 彬
(河南科技大学食品与生物工程学院/ 食品微生物河南省工程技术研究中心,河南 洛阳 471023)
由禽多杀性巴氏杆菌(Pasteurella multocida,Pm)病引起的禽霍乱是一种接触性传染病,呈世界性分布,在我国依然是常见的禽类多发病之一,给养禽业带来了一定的经济损失。对于该病的治疗主要采用抗生素,虽然效果较好,但也存在产生耐药性风险及药物残留等问题,影响产蛋率。对于该病的预防,目前临床上常用的疫苗包括弱毒活疫苗和灭活疫苗两种,其中弱毒活疫苗是我国禽疫苗中研究较多的一类。但Pm 抗原结构较为复杂,且易发生变异,导致目前商品化弱毒疫苗的免疫效果并不是特别理想。现有的市售弱毒苗存在免疫期短、保护力低、副作用偏大等问题。而灭活苗的免疫效果尚不及弱毒疫苗,因此有必要研制新型有效的疫苗。包括重组亚单位疫苗、DNA 疫苗、缺失突变活疫苗等在内的多杀性巴氏杆菌病新型疫苗均有相关研究报道[1-5]。其中DNA 疫苗因具有制备方便、成本低廉、可同时诱导产生体液和细胞免疫应答等优点,已经成为疫苗学领域研究的热点之一[6]。本实验室前期制备了PmptfA基因的裸DNA 疫苗和壳聚糖纳米DNA 疫苗,虽然均可为实验动物提供一定的保护,但仍不及弱毒活疫苗[7]。因此,需采取措施进一步提高其免疫效果。
本研究在前期研究的基础上由牡丹籽粕中提取蛋白,酶解后获得酶解产物,以其为佐剂与PmptfA基因的DNA 疫苗混合后免疫动物,并对其诱导的体液和细胞免疫应答水平及攻毒保护效果进行了检测,为Pm 病DNA 疫苗及新型疫苗佐剂的研究提供一定的参考。
1 材料与方法
1.1 主要实验材料 Pm 强毒株CVCC474 购自中国兽药监察所。Pm 弱毒活疫苗为齐鲁动物保健品公司产品。1日龄雏鸡购自河南科技大学医学技术与工程学院。限制性内切酶、T4 DNA 连接酶购自NEB 公司;HRP 标记的兔抗鸡IgG 购自北京鼎国昌盛生物技术公司;鸡IL-2、IL-4 检测试剂盒为嘉美生物技术有限公司产品;菠萝蛋白酶为Sigma 公司产品;牡丹籽粕由本实验室保存;重组蛋白PtfA(rPtfA)由本实验室制备。
1.2ptfA基因重组质粒的构建 常规方法提取Pm CVCC474 菌株的基因组DNA (引物略),以其为模板,PCR 扩增ptfA基因。扩增产物经测序验证,经BamHⅠ/EcoRⅠ酶切后与真核表达载体pcDNA3.1(+)连接,转化大肠杆菌感受态细胞JM83,提取质粒酶切鉴定后获得含ptfA基因的重组质粒,命名为pcP。
1.3 牡丹籽蛋白的提取 采用碱提酸沉法提取牡丹籽蛋白[8]。将牡丹籽粕粉碎后按照1∶25 的比例加入蒸馏水,以NaOH 调节pH 值为9.25,53 ℃水浴加热68 min 后高速离心取上清,以HCL 调节pH 值为3.5,高速离心,将沉淀冷冻干燥后获得牡丹籽蛋白粉,以考马斯亮蓝法测定蛋白含量,冻存备用。
1.4 牡丹籽蛋白酶解产物的制备 将牡丹籽蛋白粉溶于去离子水,置于80 ℃水浴中预热20 min 后调节pH 为6.5,加入菠萝蛋白酶,48 ℃水浴作用4 h,置于沸水浴灭活10 min,以冰浴冷却后10 000 r/min离心10 min,上清液经真空浓缩,冷冻干燥后获得牡丹籽蛋白酶解产物,-20 ℃保存备用。
1.5 动物免疫 大量制备ptfA基因的重组质粒pcP及空载体pcDNA3.1(+)。将120 只1日龄雏鸡饲养至6 周龄时随机平均分为6 组,分别为DNA 疫苗组、佐剂-DNA 疫苗组、灌胃组、弱毒疫苗组、pcDNA3.1(+)组和PBS 组。灌胃组将冻干的牡丹籽酶解产物以生理盐水配制成5 mg/mL 的溶液进行灌胃,每只鸡每天灌胃0.2 mL,连续灌胃15 d,期间其它各组正常饲养,灌胃结束后对各组实验鸡同时进行疫苗免疫,免疫方式为股四头肌注射。DNA 疫苗组和灌胃组将pcP 质粒调整浓度为1 μg/μL 后进行免疫,每只鸡免疫200 μL。佐剂-DNA 疫苗组向pcP 质粒中添加冻干的牡丹籽酶解产物使其终浓度为25 %且调整DNA 含量为 1 μg/μL,随后每只鸡注射200 μL。pcDNA3.1(+)组的鸡每只注射浓度为1 μg/μL 的空载体 200 μL。PBS 对照组鸡每只注射200 μL 1×PBS (pH7.2)。以上各组均免疫 3 次,每次间隔2 周。弱毒活疫苗组仅于初次免时肌注1 羽份/鸡。
1.6 免疫鸡抗体检测 初次免疫后每周采血,分离血清,分别以纯化的rPtfA 蛋白和Pm 全菌体为包被抗原,以待检血清(1 ∶100)为一抗,以兔抗鸡IgG-HRP (1∶2 000)为二抗,进行间接ELISA 试验,在OD490nm处测定吸收值以确定各组免疫鸡的血清特异性抗体水平,共检测6 周。
1.7 淋巴细胞增殖水平测定 各免疫组鸡分别于每次免疫后2 周时无菌心脏采血,分离外周血淋巴细胞,细胞计数后以RPMI1640 培养基调整浓度为1×107个/mL。96 孔细胞培养板每孔加入50 μL 细胞悬液,同时设阴性对照。实验孔和阴性对照孔各设3 个重复,实验 孔每孔加入 50 μL 10 μg/mL 的ConA,阴性对照孔每孔加入50 μL RPIM1640 培养液,置37 ℃,5%CO2培养60 h 后每孔加入5 mg/mL的MTT 10 μL,继续培养3 h。然后每孔加100 μL SDS-HCL,继续作用2 h 终止反应,测定OD570nm吸光值,计算刺激值SI=OD570nm(实验孔)/OD570nm(阴性对照孔)。
1.8 细胞因子分泌水平的测定 每次免疫2 周后,采血分离外周血淋巴细胞,调整浓度为1×107个/mL。按上述同样方法制备ConA 诱导的外周血淋巴细胞,37 ℃ 5 % CO2培养60 h 后,吸取培养上清离心收集后-20 ℃保存。采用相应检测试剂盒对各组免疫鸡分泌的IL-2 和IL-4 水平进行检测。
1.9 动物攻毒试验 第3 次免疫2 周后,对各免疫组的鸡以5LD50/ 只肌肉注射Pm 强毒菌株CVCC474进行攻毒。攻毒后继续饲养观察15 d,记录各组实验动物的死亡数量,评价各疫苗的保护效果。
2 结 果
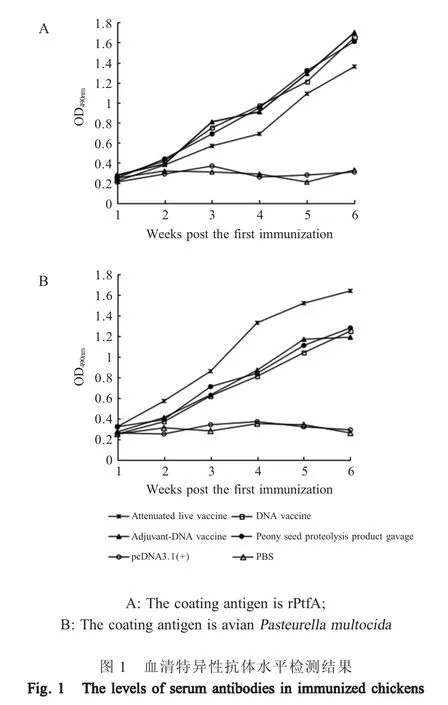
2.1 血清抗体水平检测结果 初次免疫后每周采血,采用ELISA 方法测定各组免疫鸡的特异性抗体水平,结果显示。以rPtfA 蛋白为包被抗原时,检测到的DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组免疫鸡的血清抗体水平明显高于弱毒活疫苗组(p<0.05)。而以全菌体为包被抗原时检测到的抗体水平则反之,弱毒活疫苗组显著高于上述3 组(p<0.05)。但无论以rPtfA 蛋白还是全菌体为包被抗原,DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组鸡的血清抗体水平之间则无明显差异(p>0.05) (图1)。表明牡丹籽蛋白酶解产物作为佐剂或预先灌胃对ptfA 基因DNA疫苗诱导的血清抗体水平均无明显促进作用。
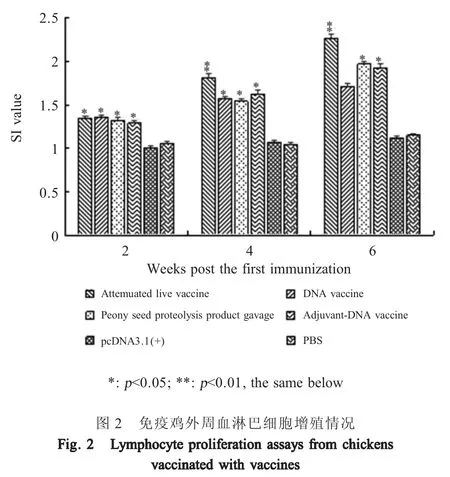
2.2 外周血淋巴细胞增殖试验结果 每次免疫2 周后,各组免疫鸡采血并分离制备外周血淋巴细胞悬液,以ConA 刺激后,MTT 法检测其增殖情况。结果显示,初次免疫后各疫苗组鸡之间的SI 值无明显差异(p>0.05)。二免后弱毒疫苗组 SI 值明显高于DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组(p<0.05),且极显著高于pcDNA3.1(+)空载体组和PBS 组(p<0.01),但DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组之间的SI 值则差异不显著(p>0.05)。第3 次免疫后,弱毒疫苗组的SI 值仍显著高于佐剂-DNA 疫苗组和灌胃组(p<0.05),且极显著高于 DNA 疫苗组(p<0.01),佐剂-DNA 疫苗组和灌胃组之间无显著差异(p>0.05),但均显著高于DNA 疫苗组(p<0.05) (图2)。表明牡丹籽蛋白酶解产物可在一定程度上提高ptfA 基因DNA 疫苗诱导的免疫鸡外周血淋巴细胞的增殖水平。
2.3 细胞因子检测结果 每次免疫后2 周时制备ConA 刺激的免疫鸡外周血淋巴细胞悬液,测定其分泌的IL-2 和IL-4 水平,结果显示。初次免疫后各疫苗组鸡外周血淋巴细胞分泌的IL-2 和IL-4 水平均无显著差异(p>0.05)。2 免和 3 免后,佐剂 -DNA 疫苗组和灌胃组鸡分泌的IL-2 的量显著高于DNA 疫苗组(p<0.05),但低于弱毒疫苗组(p<0.05),且弱毒疫苗组极显著高于DNA 疫苗组(p<0.05)。2 免后,DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组分泌的IL-4 水平无显著差异(p>0.05),但均低于弱毒疫苗组(p<0.05)。3 免后弱毒疫苗组鸡的IL-4 水平显著高于佐剂-DNA 疫苗组和灌胃组(p<0.05),且极显著高于DNA 疫苗组(p<0.01),佐剂 -DNA 疫苗组鸡虽比灌胃组鸡分泌IL-4 的量稍高,但两者无显著差异(p>0.05),且均明显高于 DNA 疫苗组(p<0.05) (图3)。表明牡丹籽蛋白酶解产物作为佐剂和预先灌胃对ptfA 基因DNA 疫苗诱导免疫鸡外周血淋巴细胞分泌IL-2 和IL-4 的能力均有一定的增强作用。

2.4 动物攻毒试验结果 3 免2 周后攻毒,连续观察15 d,记录各组鸡的存活情况,结果显示。pcDNA3.1(+)和PBS 对照组的鸡分别在攻毒后第5 d 和第6 d 全部死亡。弱毒疫苗组的鸡在攻毒后第4 d开始出现死亡,第5 d 后存活数保持不变。DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组均在攻毒后第3 d开始有鸡死亡,在第7 d~8 d 后不再出现死亡鸡(图4)。攻毒15 d 后计算弱毒疫苗组、DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组的保护率分别为85%、55%、70 %和60 %。表明牡丹籽蛋白酶解产物作为佐剂和预先灌胃均可提高ptfA 基因DNA 疫苗的保护效果,且以其作为佐剂时效果更优,但其提供的保护效果均不如弱毒疫苗组。

3 讨 论
禽霍乱是影响禽类健康的重要疫病之一,现有传统疫苗存在许多不足之处,因此研制新型疫苗势在必行。本研究室前期以Pm 的ompH、ompA和ptfA基因构建了相应的DNA 疫苗,但动物实验结果显示上述DNA 疫苗的保护效果均不及弱毒活疫苗,以ompH和ompA基因融合的DNA 疫苗的免疫效果虽优于单价DNA 疫苗,但也仅与弱毒疫苗相当[9]。因此,需采取措施如使用新型疫苗佐剂等进一步提高Pm DNA 疫苗的免疫效果。
生物蛋白经蛋白酶解后的主要产物为多肽,许多生物活性肽具有较好的保健功能,如抗氧化肽、降压肽、免疫活性肽等。免疫活性肽是一类存在于生物体内具有免疫功能的多肽,在体内一般含量较低,结构多样,是一种细胞信号传递物质,可通过内分泌、旁分泌等多种方式行使其生物学功能,增强机体的免疫功能。免疫活性肽的相关研究已成为免疫学领域研究的热点之一,包括大豆多肽、花生多肽、小米多肽等在内的多种多肽的免疫调节作用均有相关研究[10-13]。且目前也有研究显示某些多肽可增强DNA 疫苗的免疫效果[14-15]。
牡丹属于芍药科芍药属多年生木本植物,是我国的传统名花之一,不仅具有极高的观赏价值,还具有一定的药用和食用价值。牡丹籽作为生产丹皮的副产物,其油脂含量较高,目前主要用于油料加工的原料,牡丹籽榨油后的籽粕中含有丰富的蛋白,含量可达20 %~30 %[16-17]。因此有必要对其中的蛋白尤其是其酶解后的产物进行进一步的研究和利用。
本实验提取牡丹籽蛋白并以菠萝蛋白酶进行了酶解,研究了酶解产物对PmptfA基因DNA 疫苗在鸡体内免疫效果的影响。抗体检测结果显示以rPtfA蛋白和Pm 全菌体为包被抗原时,灌胃组和佐剂-DNA 疫苗组均不能提高ptfA 基因DNA 疫苗诱导产生的血清抗体水平。以rPtfA 蛋白为包被抗原时检测到的DNA 疫苗组、佐剂-DNA 疫苗组和灌胃组的血清抗体水平高于弱毒疫苗组,而以全菌体为包被抗原时检测结果则反之,其原因应与疫苗所含抗原成分有关,当以rPtfA 蛋白包被时,包被抗原成分与DNA 疫苗相似,而弱毒疫苗免疫动物后机体会产生针对多种抗原的抗体,因此检测到的针对单一抗原的抗体水平较低,低于其它三组,反之以全菌体包被时亦是如此。淋巴细胞增殖试验和细胞因子分泌水平的测定结果则显示牡丹籽蛋白酶解产物通过预先灌胃或作为佐剂均对ptfA 基因DNA 疫苗诱导的淋巴细胞增殖水平和IL-2、IL-4 分泌水平均具有一定的促进作用,表明其对该DNA 疫苗诱导的细胞免疫应答具有一定的增强作用。攻毒保护率是评价疫苗免疫效果的直观指标之一,本实验结果表明,灌胃牡丹籽蛋白酶解产物或以其为佐剂均可在一定程度上提高PmptfA基因DNA 疫苗为实验鸡提供的保护率,尤以佐剂-DNA 疫苗组效果较好,但仍达不到弱毒疫苗的保护水平。为进一步提高其免疫效果,后续实验可通过调整牡丹籽蛋白酶解产物与DNA 疫苗的比例,或对酶解产物中的免疫活性肽纯化和结构鉴定后以其为佐剂研究其效应,也可尝试以其它蛋白酶或多种蛋白酶联合酶解牡丹籽蛋白后研究其作为DNA 疫苗佐剂的效果。
