林芝地区光核桃种子表型性状变异的影响因素分析
2019-11-13刘会芳关法春吴玉德宗宪春桑利群
刘会芳,鹿 鑫,关法春,吴玉德,宗宪春,桑利群
(1.佳木斯大学生命科学学院,黑龙江 佳木斯 154007;2.牡丹江师范学院生命科学与技术学院,黑龙江 牡丹江 157012;3.吉林省农业科学院,吉林 长春 130033;4.西藏农牧学院,西藏 林芝 860000)
光核桃(Prunus mira koehne Kov et.Kpst)属于被子植物门木兰纲蔷薇科乔木植物[1],其生境多在pH 6.5 ~7.6 的砂壤上[2-3],生命过程甚至可高达上千年[4],主要分布区域在海拔2 500 ~3 600 m 之间的藏东南地区[5].目前国内外关于光核桃的研究多停留在其资源量、品质分析等[6-7]方面上,而对光核桃种子的基础生物学方面研究仍然是热点问题[8].
表型性状的遗传变异情况是评价遗传变异的简单有效方法,可全面的揭示种源间的遗传变异规律以及植物个体对于不同环境相应的适应水平,因此国内外对关于表型遗传多样性的研究一直都比较重视.光核桃在桃属中的地位相对比较原始[9],但其在西藏分布的光核桃遗传多样性程度较高,而且在其花粉表型性状上表现出很高的变异性[10],这为后续开展光核桃种质资源多样性研究奠定了强有力的理论基础.近年来,西藏地区因其地理分隔、资源破坏等不利条件的限制以及乱砍滥伐等因素影响,造成光核桃面临濒危状况[11-12].因此对其遗传多样性特点及其影响因素的研究迫在眉睫,这可以为揭示物种的进化潜力、遗传资源保护与可持续开发提供参考[13-14].
本研究以西藏林芝地区的光核桃为研究材料,主要对光核桃种子各表型性状指标进行相应的测量分析,以期通过对光核桃种子表型性状变异特点的分析和对其产生影响的相关因素进行探究,来揭示地理因素变化与遗传变异之间的关系及果实品质的影响因素,为光核桃遗传资源的保护提供理论依据.
1 材料与方法
1.1 材料和测定方法
光核桃种子采自林芝市巴宜区、波密县、朗县等地,在9 月末光核桃果实成熟季节,每个样点随机选择光核桃果树,并摘取10 粒无畸形无病的果实,将果实充分后熟半个月后,再从中随机选取3 粒果实,取出种子,用游标卡尺测量样本横径、纵径、厚度等指标,3 次重复,并计算相应的果形指数(果形指数=纵径÷横径).具体采样地点见图1.

图1 采样点分布地图Fig.1 The map of point distribution
1.2 数据统计分析方法
使用SPSS 20.0 软件分析对种子各表型性状指标数据进行分析;对各光核桃种子样本进行聚类分析得到聚类图;同时对光核桃种子表型性状测量值与其相对应的海拔高度进行Pearson 相关系数分析.
2 结 果
2.1 光核桃种子表型性状描述及特征分析
由表1 来看,种子的横径数值大小分布在10.95~19.53 之间,变异系数为11.70%;纵径的数值大小分布在10.90 ~26.17 mm;变异系数为12.13%;种子的厚度数值大小分布在7.83 ~13.41 mm 之间,变异系数为10.43%;单粒重的数值大小分布在0.43 ~1.49之间,变异系数为28.71%.因此,相对而言,单粒重的变异系数最大,则说明其的变异幅度较大,受环境因子的影响较大;种子厚度变异系数最小,则说明其受环境的影响较小,该性状是相对较稳定的;各表型性状的P 值均小于0.01,则表明光核桃种子的表型性状存在极显著的遗传变异.

表1 光核桃种子表型性状的方差分析Table 1 Variance analysis of phenotypic traits of Prunus mira seeds
2.2 表型性状的聚类分析
采用光核桃表型性状指标计算各样本间欧氏距离,并进行聚类分析.50 组光核桃种子可分为3 个类群(图2),第1 类群包含15 组样本,P36 ~P50,占总样本的30%;第2 类群包含2 组样本,分别是P11、P35,占总样本的4%;第3 类群包含剩余33 组样本,P1 ~P10、P12 ~P34,占总样本的66%.其中,第2 类群和第3 类群在欧氏距离为14 阈值时分别可以聚成一类,则说明其亲缘关系较近.
第1 类群来自于朗县,米林县和巴宜区的样本14组,来自于工布江达县的有1 组;第2 类群第一组样本来自波密地区,第二组样本来自朗县,米林县和巴宜区;第3 类群19 组样本来自于波密地区;来自于朗县,米林县和巴宜区的样本11 组,来自于工布江达县的样本有3 组.结合采样地点分布图得知,不同样本的形态分类与地理来源并无严格的一致关系.
2.3 光核桃种子表型性状和可溶性固形物的相关性分析
由表2 可知,在种子的主要表型性状中,相关性主要表现在横径与纵径、厚度、单粒重,厚度与纵径、单粒重,纵径与果形指数和单粒重,可溶性固形物与单粒重呈极显著正相关;果形指数与可溶性固形物呈显著正相关;横径与果形指数,厚度与果形指数呈极显著负相关.其他指标之间相关性较小.
2.4 光核桃种子表型性状、可溶性固形物含量与海拔的相关性分析
由表3 可知,在光核桃种子表型性状、可溶性固形物含量与海拔的相关性分析中,相关性主要表现在海拔高度与纵径、厚度和单粒重呈极显著负相关,与果形指数表现为显著负相关;与横径和可溶性固形物含量的相关性较小.西藏光核桃群体间存在基于海拔梯度上的遗传变异,具有明显的地理区域特性,但海拔因素对光核桃的果实品质影响不大.
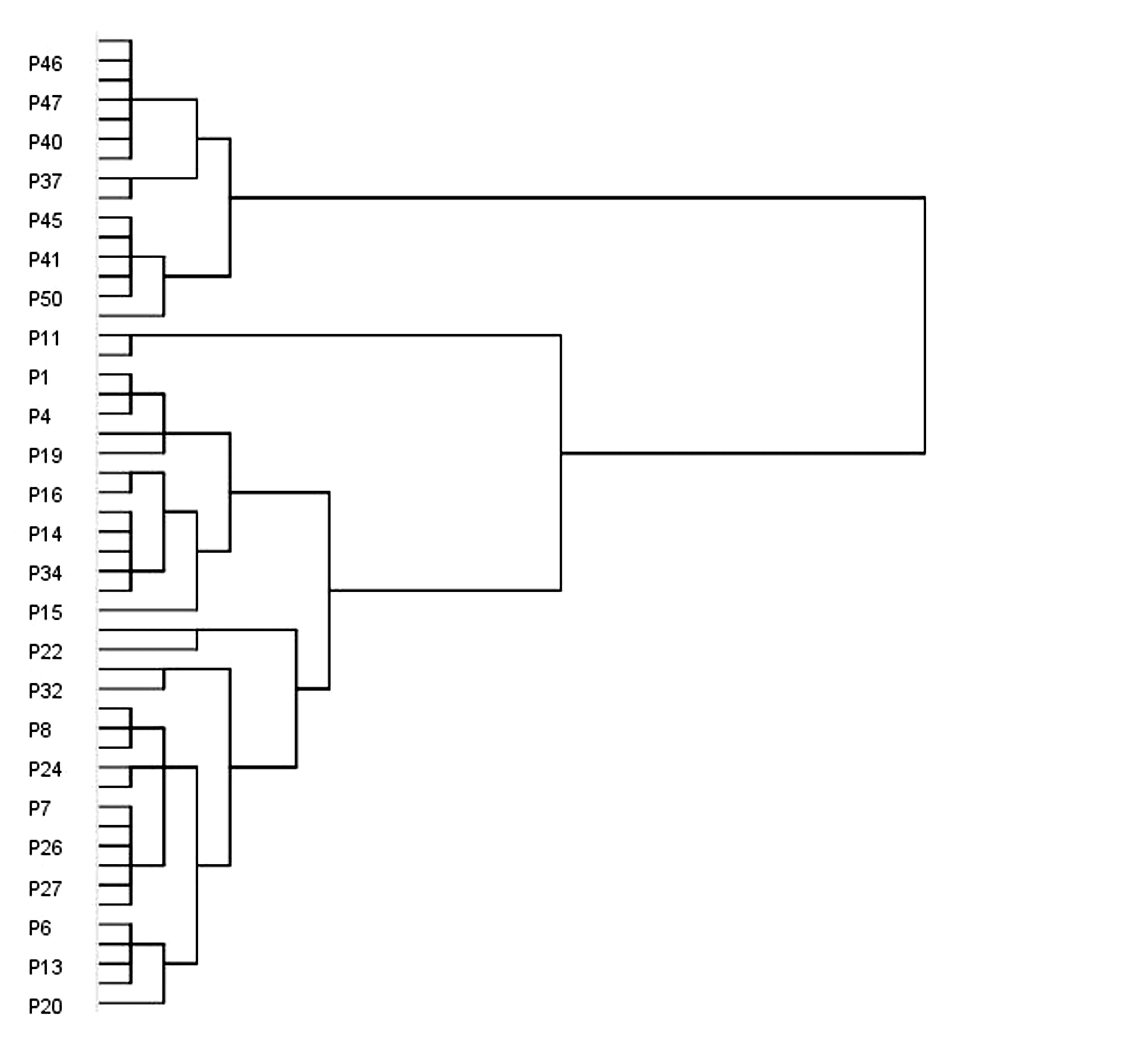
图2 表型性状的聚类分析图Fig.2 Cluster analysis of phenotypic traits
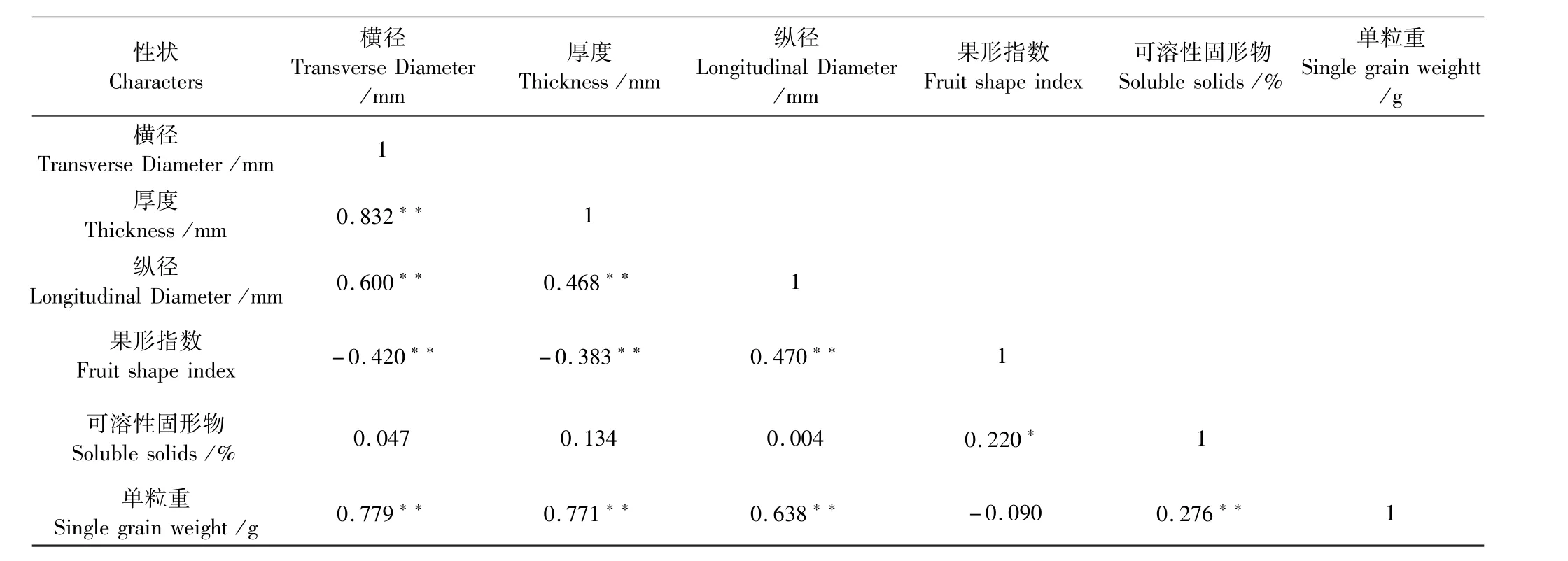
表2 光核桃种子表型性状和可溶性固形物的相关性分析Table 2 Correlation analysis of phenotypic characters and soluble solids of Prunus mira seeds

表3 光核桃种子表型指标、可溶性固形物含量与海拔高度相关性分析Table 3 correlation analysis of Prunus mira seed phenotype index,soluble solid content and altitude
3 讨 论
3.1 光核桃种子表型性状的遗传变异与性状相关
表型性状研究是生物遗传多样性分析的方法之一[15].植物个体表型性状是基因与环境交互作用的结果,物种在长期的进化过程中也会为适应各种不同的环境,产生与环境相适应的包括表型性状在内的遗传性状[16],并在后代中将稳定的性状表现出来.不同地域光核桃种子的横径、纵径、厚度等性状之间存在一定程度上的差异,其中种子单粒重变异系数最大,性状变异幅度较大,而种子厚度变异系数最小,性状变异较小,则说明该性状是相对稳定的;种子纵径及横径的变异系数则居中;光核桃的各表型性状的P 值均小于0.01,则意味着光核桃种子的表型性状存在显著的遗传变异.基因和环境都可能是导致这种情况的因素,这在其他以往高原地区同类研究中也得到类似结果[17].
可溶性固形物含量是衡量果实果肉品质的重要指标[18].光核桃种子横径与其他表型性状相关性最高,而果实可溶性固形物含量与单粒重呈极显著正相关,与果形指数呈显著正相关关系,与其他表型性状间相关关系较小.光核桃大部分表型性状相关性相对较高,这与以往结果研究相似[19],这其中蕴含的性状连锁之间的关系,今后可通过通径分析等方法进一步深入研究.
3.2 光核桃种子表型性状聚类分析及与海拔相关性
对植物种质性状进行聚类分析常用来研究多样化群体亲缘遗传关系[20].对50 组光核桃种子表型性状指标的聚类分析可分成5 个大的类群,结合采样地点的地理位置分析,各大类群各种质资源亲缘关系与该样本的来源地无必然联系.因此,地理区隔因素不是光核桃在表型性状上产生差异的主要原因.海拔高度与光核桃种子纵径、厚度、果形指数、单粒重等指标具有高度负相关关系,海拔高度是影响林芝地区光核桃表型性状生物进化和遗传变异的一个关键性因素.随着海拔的升高,生存环境恶劣、基因流动困难,气候和植被等也呈现明显的垂直变化[21].不同海拔居群所处生境的温度、降雨量、光照等生态因子均存在差异,导致不同海拔种群植物生境存在差别,从而影响植物的生长和发育过程,最终导致种群间在遗传性状等方面产生明显的遗传分化现象[8,22-23].
4 结 论
各表型性状P 值均小于0.01,说明光核桃种子的表型性状产生了显著的遗传分化,其中,单粒重的变异系数最高(28.71%),厚度的变异系数最低(10.43%);聚类方法分析后可将不同样本种质资源整体分为3 个类群,但不同样本的形态分类与地理来源无一致关系,而海拔高度与光核桃种子的各表型性状高度相关,同时果形指数及单粒重与果肉可溶性固形物高度相关.因此,西藏林芝地区光核桃在表型性状上存在由海拔梯度引起的变异,而非地理区隔因素.
