水杨酸和茉莉酸诱导水稻化感关键基因C4H的表达分析
2019-11-11刘亚洲孙一丁郭怡卿
刘亚洲 ,张 翠,孙一丁*,杨 奕,李 铷,卢 红,郭怡卿*
(1.云南农业大学 植物保护学院,云南 昆明 650201;2.云南省农业科学院生物技术与种质资源研究所,云南 昆明 650223)
【研究意义】“化感作用”(Allelopathy)是指植物(包括微生物)通过其本身产生并释放到周围环境中的化学物质对另一种植物(或微生物)产生直接或间接的相互排斥或促进的效应,是对环境的一种适应机制和对逆境的防御机制[1]。提高农作物的抗逆性是高产稳产的重要基础,也是减少化学品投入、保护生态环境的重要保障。作物化感抗性品种的选育可提高其自身种质的抗逆性、提高生物防治效率,是减少化学品投入、保护生态环境的根本措施[2]。以减少产量损失为目标的抗性育种(抗逆境、抗病虫害等)是水稻育种的重要目标之一。水稻化感抗性为这一回标的实现提供了新的途径。【前人研究进展】水稻化感的研究始于20世纪80年代中后期,美国科学家Dilday等对种质资源库所收集的12 000份水稻进行了抑草评价,发现大约3 %的种质材料具有化感抗杂草潜力[3-6]。水稻化感现象引起各国科学家的兴趣,纷纷开展了水稻化感抗性种质评价、化感物质分离与鉴定、抗性遗传性等方面的研究并取得重要进展[7-9]。相关研究表明,酚酸类化合物是重要的化感物质[1-4]。与非化感水稻相比,化感水稻能够通过其分泌的酚酸类化感物质介导而影响根际微生物活性,同时抑制杂草生长;逆境引起的水稻化感作用增强,导致了合成酚酸类物质的基因表达增强以及释放到根际土壤中的该类物质量增多[11-13]。苯丙烷途径是植物化感物质代谢合成中的重要途径,肉桂酸-4-羟化酶(C4H)又是酚酸类物质代谢的关键酶[14]。C4H在植物中多以小基因家族的形式存在,不同物种间基因数量不同[15],以应对植物体复杂的生理调控[16]。水杨酸(SA)和茉莉酸(JA)作为重要的化学信号,能激活很多防卫反应[17-19],内源与外源的SA、JA均有诱导植物的获得性抗性的作用,如水稻抗病性[20]以及耐冷与抗旱等生物和非生物胁迫抗性[21-23]。何海斌等通过MeSA和MeJA处理化感水稻IAC165和非化感水稻华粳籼,发现2种水稻的化感潜力得到增强,酚酸类化合物分泌增多,相关酶活性也发生改变[24]。【本研究切入点】水稻中有4个编码C4H的P450基因[25],分别是Os01g0820000(CYP73A35),Os02g0467000(CYP73A39),Os02g0467600(CYP73A40)和Os05g0320700(CYP73A38),在化感与非化感水稻材料中这些基因的表达是否存在差异及其与化感潜力的关系,是明确水稻化感抗性机制的重要基础。【拟解决的关键问题】本研究以化感水稻PI312777及非化感水稻Lemont为研究对象,采用SA和JA处理,q-PCR(Quantitative real-time Polymerase Chain Reaction,简称q-PCR)分析4个C4H基因的表达差异,同时通过水浸提液生物测定的方法测定2种水稻化感潜力的变化,以期望发现这4个基因与水稻化感潜力的相关性,揭示酚酸类物质与水稻化感潜力发挥的内在分子机制。
1 材料与方法
1.1 供试材料
选用化感水稻PI312777(PI)和非化感水稻Lemont(Le)[3]作为供体植物,以稗草作为受体植物。稗草种子为上一年度田间采集。同时配制浓度为100 mM的SA,浓度为200 mM的JA备用。
1.2 实验设计
化感水稻PI312777(PI)和非化感水稻Lemont(Le)分别用JA和SA处理,其中,处理Le(CK),未经处理的Lemont;处理Le(JA),经过茉莉酸处理的Lemont;处理Le(SA),经过水杨酸处理的Lemont;处理PI(CK),未经处理的PI312777;处理PI(JA),经过茉莉酸处理的PI312777;处理PI(SA),经过水杨酸处理的PI312777。
1.3 实时荧光定量分析
育苗及前处理:在育苗穴盘中填充育苗基质,分别播种化感与非化感水稻材料,在温室中生长到3~4叶期,分别将100 mM的SA和200 mM的JA用喷壶均匀喷洒到2种水稻的叶面上,分别放置于光照培养箱中,25 ℃,12 h光/暗培养。在处理后1、3、24 h分别剪取水稻叶片并立即放入液氮,随后置于-80 ℃保存备用。
RNA提取:已保存的不同处理水稻叶片采用普洛麦格试剂盒(Eastep®Super)提取RNA,按说明书操作,并采用微量紫外分光光度仪测量RNA的含量。
反转录:上述提取的RNA采用普洛麦格(GoScriptTMReverse Transcription System)第一链合成试剂盒20 μl体系进行反转录获得cDNA。
引物设计:用肌动蛋白β-Actin作为内参引物,同时在NCBI上设计4个基因的扩增引物见表1,由北京硕擎生物公司昆明分部合成,ABI QuantStudio 6 Flex荧光定量PCR仪进行q-PCR分析各基因的相对表达量。
q-PCR分析:采用上海生工SGExcel FastSYBR Mlxture(With ROX)20 μl体系。即上游引物0.4 μl,下游引物0.4 μl,Mix 10 μl,RNase-free ddH2O 8.2 μl,模板RNA 1 μl。q-PCR 程序设定为 95 ℃预变性3 min,95 ℃变性15 s,60 ℃退火延伸20 s,45个循环;95 ℃变性15 s,60 ℃ 1 min,95 ℃变性15 s,60 ℃ 15 s,基因相对表达量按照 2-ΔΔCt法计算,基因的相对表达量=2-ΔΔCt,其中ΔΔCt=ΔCt(未胁迫)-ΔCt(胁迫),ΔCt(样本)=Ct(目标基因)-Ct(参照基因)。

表1 4个C4H基因q-PCR引物
1.4 化感潜力分析
采用叶片水浸提液[24-25]方法进行,水浸提液是一种常用的评估水稻化感潜力的室内生物测试法。分别取经SA、JA处理48 h后相同部位的水稻叶片0.5 g,剪成0.2 cm左右大小的小段,加入5 mL灭菌蒸馏水4 ℃下浸提24 h,用灭菌过滤器滤出获得水浸提液。将稗草种子用质量分数为6 %次氯酸钠表面消毒,灭菌水清洗后均匀播种在培养皿中,萌发至露白,转移到含有2 %的琼脂培养基上,根部向下,每皿20粒,封盖后将该培养皿放置在25 ℃的光照培养箱内,12 h光/暗垂直培养2 d,选留主根长0.5~1 cm的植株,将上述获得的水浸提液每皿10株,用微量移液器分别处理稗草的根尖部分,每植株10 μl,待处理部位干后将培养皿再次封盖放回培养箱培育3 d,测量稗草的主根长度,计算根长抑制率IR评价化感潜力变化。试验在无菌条件下进行操作,最大限度地控制了微生物的干扰和微生物对化合物的分解直接处理受体植物生长点(根尖),使试验所需要的水浸提液的量减少,适合于试验材料稀少的情况采用。IR的计算公式如下:
IR= (1-Tr/ck) × 100
式中,Tr表示处理组受体植物稗草的生长量测定值,ck表示对照组稗草的生长量测定值,IR表示供体植物水稻对受体植物稗草的影响作用,当IR<0时,表示供体植物水稻对受体植物稗草的作用为刺激生长;当IR>0时,表示供体植物水稻对受体植物稗草具有抑制作用;当IR=0时,表示供体植物水稻对受体植物稗草没有活性或表示2种植物间无化感作用存在。

图1 PI312777(PI)和Lemont(Le)在JA和SA激素处理下Os01g0820000的相对表达量
2 结果与分析
2.1 C4H基因相对表达分析
2.1.1 Os01g0820000基因相对表达量分析 JA处理后该基因相对表达量在1、3和24 h 化感和非化感水稻中均呈明显的降低趋势;SA处理后Lemont中该基因相对表达量在1和3 h降低,而24 h则出现微微的升高,PI312777中的相对表达量也与Lemont类似,但PI的表达量较Le的稍高(图1)。这一结果表明该基因在不同化感潜力的水稻中表达相似,对诱导信号具有相似的反应模式。
2.1.2 Os02g0467000基因相对表达量分析 如图2所示,JA处理后的该基因的相对表达量在2种水稻中,均会呈现出先降后升然后接近正常水平,不同的是在PI312777中,该基因在处理后的3 h相对表达量出现上调,而在Lemont中该基因则在24 h时才出现上调。但经SA处理的Lemont该基因相对表达量在1 h有较明显的升高,在3 h表现为下调,而在PI312777该基因相对表达量在1 h下调,在3 h表达量上调(图2)。试验结果显示,在化感水稻PI312777中,该基因在应对JA处理时,反应要更快。而SA处理后的2种水稻中,该基因在Lemont中反应要快于PI312777。2种水稻面对处理不同处理,均有不同的表达模式,但是结果相似。

图2 PI312777(PI)和Lemont(Le)在JA和SA激素处理下Os02g0467000的相对表达量
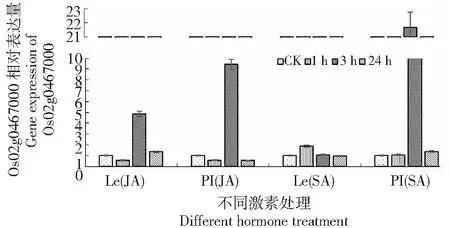
图3 PI312777(PI)和Lemont9(Le)在JA和SA激素处理下的Os02g0467600相对表达量
2.1.3 Os02g0467600基因相对表达量分析 经过JA处理后该基因相对表达量在化感和非化感水稻中均先上调后下调,且在3 h时出现较高程度的上调;而经过SA处理的该基因在2种水稻中的表达量在所测量的时间段内均出现上调,且在化感水稻PI312777中3 h时,相对表达量上调幅度较大。从该基因相对表达量可以看出来,面对SA和JA的胁迫,2种水稻的该基因均表现出一定的上调以应对外源激素的胁迫,不同的是化感水稻PI312777的该基因的相对表达要高于非化感水稻Lemont,说明该基因在PI312777中面临胁迫时,更多的是参与到了肉桂酸-4-羟化酶的合成(图3)。
2.1.4 Os05g0320700基因相对表达量分析 经过JA和SA处理后,非化感水稻Lemont在1、3和24 h时该基因的相对表达量均表现出下调。而经过JA和SA处理后的化感水稻PI312777的该基因相对表达量均出现先下调后上调的情况,不同的是经过JA处理的PI312777在3 h表达量出现上调,而经过SA处理的该基因相对表达量在24 h才出现上调(图4)。说明该基因在2种水稻中表达差异较大,在化感水稻中表达量要高于非化感水稻。
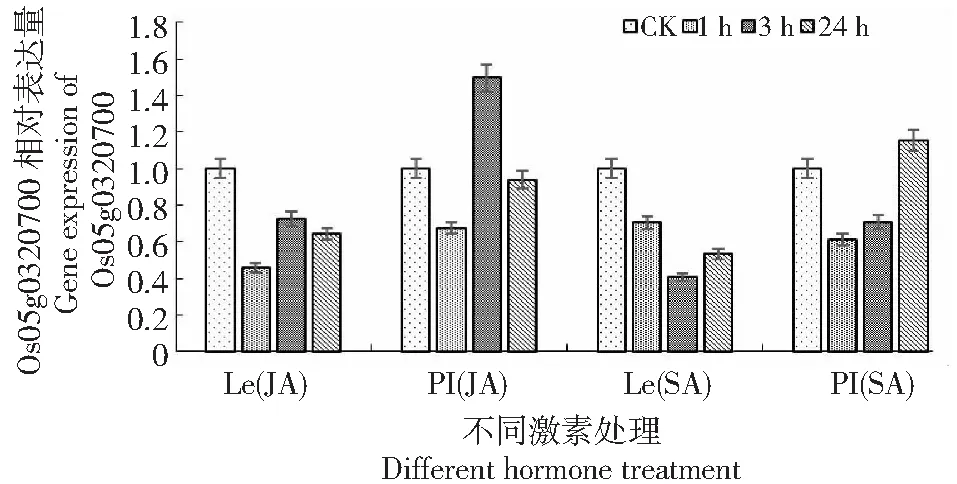
图4 PI312777(PI)和Lemont(Le)在JA和SA激素处理下Os05g0320700相对表达量
2.2 水浸提液化感潜力分析
JA和SA处理后水稻叶片水浸提液对稗草根生长的生物测定表明,Lemont的抑制率较未处理的分别提高了22 %和18 %;而PI312777的提高了40 %和28 %,结果表明,JA和SA处理均使2种水稻的化感潜力得到增强,JA的诱导作用高于SA,且PI312777增强均高于Lemont(表2)。
3 讨 论
茉莉酸以及水杨酸作为植物体内重要的化学信号,它们均能通过提高植物体内相关酶活性等来提高植物抗逆性[27-30]。水稻化感是对环境的一种适应机制和对逆境的防御机制,经过外源水杨酸和茉莉酸诱导后,体内相关酶活性会发生变化以应对胁迫。有研究表明,水稻在受到胁迫时,相关基因相对表达量在24 h内差异较大,而超过48 h相对表达量差异不显著;植物体内酚酸类物质在胁迫后48 h积累量达到最高[31]。因此本研究选择的时间范围在处理后24 h以内,同时还检测了处理后不同基因早期1、3 h的表达量变化。
为研究PI312777和Lemont 2种水稻的浸提液对稗草根长的影响,利用生测法对2种水稻的水浸提液进行检测。结果表明,SA、JA诱导处理提高了化感抗耐稗草的作用。而4个C4H基因在PI312777和Lemont中相对表达量存在一定差异,其中差异较大的是Os02g0467600和Os05g0320700。Os02g0467600受SA和JA诱导时,PI312777中应答更早,相对表达量的上调程度也要高于Lemont;Os05g0320700在Lemont中一直处于下调状态,而该基因相对表达量在PI312777中则均出现先下调后上调。Os01g0820000和Os02g0467000表达模式相似,差异不显著。因此推测Os02g0467600和Os05g0320700这2个基因是造成这2种水稻化感潜力不同的重要因素。

表2 水稻叶片水浸提液对稗草生长的影响
通过对模式植物拟南芥的类比分析发现[32],拟南芥中只有1个C4H基因CYP73A5,而通过突变体材料的研究发现,突变体植株的表型出现矮化,以及雄性不育、酶不稳定等症状,突变体的木质素含量明显下降,肉桂酸酯积累量增多等,说明该基因在拟南芥中受不同的转录因子调控而影响到下游基因的表达及生物体的生长发育等。通过SA和JA处理的2种水稻的化感潜力增强,但基因的表达情况却不同,说明这4个基因在水稻中分别有不同的表达模式。通过对2种水稻4个C4H基因的CDs克隆分析,发现4个基因的序列差异极小。为了揭示2种水稻化感潜力不同的原因,对化感水稻PI312777和非化感水稻Lemont启动子进行了分析,发现与非化感水稻Lemont相比,化感水稻PI312777中有多个MYB类、bHLH类、WRKY类转录因子表达量上调(数据未发表)。假设引起2种水稻化感潜力不同的原因是由转录因子引起的,因此,通过对调控化感水稻的转录因子进行克隆和功能鉴定,比较其在化感和非化感水稻中的表达情况,可进一步明确这4个基因在化感及非化感水稻中表达差异的原因,从而可进一步揭示水稻化感抗杂草的分子机制。
4 结 论
经过茉莉酸和水杨酸处理的化感水稻PI312777和非化感水稻Lemont叶片水浸提液对稗草根生长有显著的化感抑制效应的增强。本研究表明,经过水杨酸和茉莉酸处理后,肉桂酸-4-羟化酶的4个编码基因差异表达,化感的潜力变化可能与此有关,进一步研究分析发现差异表达的原因与启动子有关。
