Immunodetection of ephrin receptors in the regenerating tail of the lizard Podarcis muralis suggests stimulation of differentiation and muscle segmentation
2019-10-29LorenzoAlibardiComparativeHistolabPadovaandDepartmentofBiologyUniversityofBolognaBologna40126Italy
Lorenzo Alibardi Comparative Histolab Padova and Department of Biology,University of Bologna,Bologna 40126,Italy
ABSTRACT Ephrin receptors are the most common tyrosine kinase effectors operating during development.Ephrin receptor genes are reported to be upregulated in the regenerating tail of the Podarcis muralis lizard. Thus, in the current study, we investigated immunolocalization of ephrin receptors in the Podarcis muralis tail during regeneration.Weak immunolabelled bands for ephrin receptors were detected at 15-17 kDa, with a stronger band also detected at 60-65 kDa. Labelled cells and nuclei were seen in the basal layer of the apical wound epidermis and ependyma, two key tissues stimulating tail regeneration. Strong nuclear and cytoplasmic labelling were present in the segmental muscles of the regenerating tail, sparse blood vessels, and perichondrium of regenerating cartilage. The immunolocalization of ephrin receptors in muscle that gives rise to large portions of new tail tissue was correlated with their segmentation. This study suggests that the high localization of ephrin receptors in differentiating epidermis, ependyma,muscle, and cartilaginous cells is connected to the regulation of cell proliferation through the activation of programs for cell differentiation in the proximal regions of the regenerating tail. The lower immunolabelling of ephrin receptors in the apical blastema, where signaling proteins stimulating cell proliferation are instead present, helps maintain the continuous growth of this region.
Keywords: Lizard; Regenerating blastema; Ephrin receptors;Immunolabelling;Western blotting
INTRODUCTION
Among amniotes, only lizards regenerate a large organ such as their tail after loss (Alibardi, 2014, 2015, 2017a, 2018;Bellairs & Bryant, 1985; Cox, 1969; Fisher et al., 2012; Gilbert et al., 2015; Hughes & New, 1959; Lozito & Tuan, 2016). The process of tail regeneration in lizards occurs after the formation of a regenerative blastema over the injured tissue of the tail stump. The blastema comprises a soft mesenchyme rich in hyaluronate and is covered by an actively proliferating epidermis (Alibardi, 2017b, 2017c). While the growing tip of the blastema elongates the tail, the more proximal regions to the tail stump differentiate into axial tissues (Alibardi & Meyer-Rochow, 1989). The latter comprise a cartilaginous cylinder replacing the vertebral column and encasing the ependymal tube derived from the spinal cord and segmental myomeres of similar dimension (Figure 1). Regenerating muscles gradually grow from pro-muscle aggregates derived from the local fusion of myoblasts that give rise to elongating myotubes,while the segregated fibroblasts form the connective myosepta(Alibardi,1995;Cox,1969;Hughes&New,1959).
Recent research has revealed the expression of numerous developmental genes in the regenerating tail of lizards(Hutchins et al, 2014; Liu et al., 2015). Comparison between up- and down-regulated genes in the tail with those in limbs has helped identify key genes responsible for tail regeneration(Vitulo et al., 2017a, 2017b). Previous studies have also detected the expression sites of these genes and their coded proteins(Alibardi,2017a,2017d).Thus,based on transcriptome analysis, it is now possible to determine the localization of specific, highly up-regulated proteins responsible for tail regeneration, avoiding a random search for signaling proteins,growth factors,oncogenes,and transcription factors.
One of the more up-regulated and tail-exclusive genes was coding for a receptor of ephrin proteins (erythropoietin producing hepatocellular carcinoma receptor), ephbr6, with 8-30-fold of expression in the regenerating blastema in comparison to the tissues present in the normal tail (Vitulo et al., 2017a). Ephrins are membrane-bound proteins that stimulate their specific tyrosine-kinase receptors located on the plasma membrane of a target cell, which respond by phosphorylation of cytoplasmic proteins for specific gene activation and cytoskeletal re-organization (Klein, 2012;Palmer & Klein, 2003; Park & Lee, 2015). Ephrin receptors comprise proteins inserted in a cell membrane that interact with ephrin membrane proteins and their physiological ligands, and are distinguished as sub-class A and B. These receptors comprise the highest number of protein kinases present in cells of vertebrates, and are involved in numerous morphogenetic processes, including the development of paraxial segmentation, limb skeleton, and nervous system regionalization, as well as cancer and metastasis intervention(Durbin et al., 1998; Henkemeyer et al., 1994; Pasquale,2010; Poliakov et al., 2004). These proteins and their receptors mediate the formation of cell junctions in external or internal epithelia, during neural tube development, in growing blood vessels, during formation of temporary junctions in migrating cells of the neural crest, and establishment of compartmentalization among different tissues, thus forming the basis for the development of distinct organs (Park & Lee,2015).
In the regenerating tail of the common wall lizard (Podarcis muralis), where cell movement and tissue formation and reorganization are present, several genes coding for ephrin receptors are reported to be highly up-regulated (Vitulo et al.,2017a). However, the specific role of these genes and their main sites of expression in the regenerating tail remain undetermined. Therefore, based on immunohistochemical analysis, we detected the sites of ephrin receptor localization in the tail blastema-cone of Podarcis muralis.
MATERIALS AND METHODS
Procurement and maintenance of animals
Eight common wall lizards (Podarcis muralis), whose tail tissues were prepared earlier for previous investigations, were used in the present study. Adult females and males were captured in the wild and were then maintained in cages at fluctuating temperatures of 25-30 °C. The lizards were fed three times weekly with maggots and mealworms and exposed to natural sunlight. All animal care and handling procedures followed Italian guidelines (Art. 5, DL 116/92).After inducing autotomy at about 1/3 proximal of the tail, a natural and painless process of tail amputation, the tail was regenerated at 25-30 °C until a 3-4-mm blastema-cone formed over the tail stump (~12-16 d later). We collected tissues from three adults with 3-4-mm long regenerated tails for protein extraction and successive electrophoretic analysis.
Western blotting
Tissues were homogenized in 8 mol urea and 50 mmol Tris-HCl at pH 7.6 containing 0.1 mol 2-mercaptoethanol, 1 mmol dithiothreitol, and 1% protease inhibitor (Sigma, St Louis,USA). The non-solubilized tissue components were removed by centrifugation at 10 000 g for 5 min at room temperature.Protein concentration was assayed by the Bradford method before electrophoresis.
For electrophoresis, the same amount of protein (35 μg)was loaded in each lane and separated in a gradient gel (8-250 kDa molecular weight (MW)) using the MiniProtean III electrophoresis apparatus (Bio-Rad, USA). For Western blotting, the separated proteins were transferred to nitrocellulose membranes, which were then stained with Ponceau red to verify protein transfer and incubated with primary mouse antibody against ephrin receptor. The primary antibody (CPTC-EPHB4-1) was induced in mice against an amino acid sequence within the Human Ephrin Receptor B4 protein (UniProt ID P54760). The antibody was produced by Clinical Proteomics Technologies for Cancer, National Cancer Institute, and maintained by the Hybridoma Developmental Study Bank, University of Iowa, USA, supported by the US NIH. For immunoblotting the antibody was utilized at 1:300 dilution in buffer, whereas the primary antibody was omitted in the controls. Detection of the separated and reactive protein bands was performed using enhanced chemiluminescence as per Amersham, which employed fluorescent secondary antibodies against mouse immunoglobulins (ECL, Plex Western Blotting System,GE Healthcare,UK).
Fixation and microscopic methods
The regenerating tissues were fixed for 8 h at 0-4 °C in 4%paraformaldehyde in 0.1 mol/L phosphate buffer at pH 7.4 and were then rinsed in buffer for 30 min, dehydrated in ethanol,clarified in xylene,and embedded in wax.
The tissues were sectioned (6-8 μm) using a microtome.The sections were then collected on chromolume-gelatin precoated slides and dried for 2-3 h on a warm plate.Representative sections were stained with Hematoxylin-Eosin(HE) or Mallory stain for histological examination. After dewaxing with xylene and hydration, the sections utilized for immunofluorescence were initially treated for antigen retrieval using a microwave-oven technique. The sections were immersed in 0.1 mol/L citrate buffer at pH 5.6, and tissues were exposed to irradiation for 6 min.After this, sections were rinsed in Tris buffer 0.05 mol/L at pH 7.6 containing 5% bovine serum albumin and were pre-incubated for 20 min with buffer containing 2% normal goat serum to block non-specific antigens. The sections were incubated for 6 h at room temperature with primary mouse antibody (see above) at 1:50 dilution in buffer, whereas control sections were incubated with buffer only (omitting primary antibody). After rinsing in buffer for 10 min three times, the sections were incubated for 60 min at room temperature with fluorescein-isothiocyanate conjugated to an anti-mouse IgG (FITC, Sigma, USA), diluted 1: 200 in buffer. The sections were again rinsed three times in buffer, mounted in anti-fading medium (Fluoroshield, Sigma,USA), and observed under a fluorescence microscope using a fluorescein filter. Pictures were taken using a digital camera and digitalized using the Adobe Photoshop Program v8.
RESULTS
Bioinformatics and Western blotting
Bioinformatics analysis using the Clustal-W Muscle Program showed that at least three ephrin-receptor proteins in the lizard Anolis carolinensis possessed possible epitopes recognized by the antibody employed here; in particular,ephrin receptor-B4 showed a higher identity in comparison to ephrin receptor-B2 and-B3(Figure 2).
Immunoblot detection showed a main band at 60-65 kDa and weak bands at 130-140 and 17 kDa (Figure 3, first lane).Control blots showed lower labelling only at ~65 kDa,suggesting that this band also contained non-specific protein material packed with reactive antigen(Figure 3,second line).
Histology
The regeneratin g tail was composed of mesenchymal and loose connective tissues at the tip for 0.3-0.5 mm. This apical tissues were in continuation with a loose connective containing fibroblast-like cells in more proximal regions 0.5-1.0 mm from the tip (Figure 4A, B).The thick wound epidermis of the apical blastema consisted of numerous layers of keratinocytes covered by a thin corneous layer (Figure 4B).The apical epidermis was undulated and formed one or more small apical pegs. Numerous melanocytes were seen in the blastema and they also infiltrated the basal layer of the regenerating epidermis. Near the apical epidermis, a tube of ependymal cells, often forming a dilated ampulla, was present and surrounded by a cylinder of cartilage cells (Figure 4A).Around the cartilage, irregular loose connective tissue containing spare blood vessels and nerves was present. The inner areas of connective tissue forming the future dermis contained bundles of regenerating muscles that were degrading into small pro-muscle aggregates toward the tip of the regenerating blastema (Figures 1, 4A). Moving proximally,the muscle aggregations formed segmented units made of bundles of myotubes separated by fibroblasts that gave rise to inter-muscle connective septa in more proximal, mature regions of the regenerating tail(Figures 1,4C).Immunofluorescence

Figure 2 Epitope regions, amino acid position within protein (aa), and GenBank accession No. for three ephrin receptors detected in NIH database for Anolis carolinensis

Figure 3 Western blotting of regenerating tail showing labelled bands
The apical wound epidermis showed stronger immunolabelling in comparison to the mesenchymal blastema (Figure 5A). The numerous blood vessels fluoresced yellow, an indication of non-specific (auto-)fluorescence. Most suprabasal and external, pre-corneous keratinocytes were sparsely immunofluorescent, and the fluorescence appeared mainly localized along the peripheral cytoplasm (Figure 5B, C). Only basal cells in contact with the underlying blastema showed some nuclear labelling. Immunofluorescence decreased over the entire thickness of the proximal epidermis where scales were forming (Figures 5D, 6A). In the suprabasal layers of the regenerating scales, keratinocytes appeared in the course of differentiation to give rise to the spindle-shaped cells of the forming corneous layer. These differentiating cells also showed some labelling along their perimeter(Figure 6A).

Figure 4 Histology of regenerating tail
The cells forming the enlarged ependymal ampulla showed variably intense immunolabelling in their cytoplasm and occasionally also in the nucleus, which appeared stronger in comparison to that seen in the mesenchymal cells of the surrounding blastema (Figure 6B, C). The differentiating cartilage cells surrounding the ependymal tube also showed more intense labelling than that in the cells of the peripheral mesenchyme (Figure 6B). Small as well as larger nerves entering the blastema also appeared more intensely labelled than the cells of the surrounding mesenchyme or of the loose connective tissue and forming dermis (Figure 6D). In more proximal regions, at 0.5-3 mm from the tail tip, the external region of the cartilaginous tube, the perichondrium, appeared intensely labelled but the labelling decreased in the fusiform chondrocytes localized in the more central regions of the cartilaginous tube (Figure 7A, B). In addition, the wall of the large blood vessels (containing few or no erythrocytes)present in the loose connective tissue surrounding the cartilaginous tube, showed high immunofluorescence (Figure 7B). Erythrocytes fluoresced intensely yellow, especially when clustered within the lumen of the blood vessels, disturbing the green fluorescence of vessel walls.

Figure 5 Immunofluorescence of blastema for ephrin receptors
The initial pro-muscle aggregates present at the apex of the regenerating blastema (Figures 1, 4A), appeared more immunofluorescent than the surrounding mesenchyme (Figure 7C). Strong immunolabelling was also noted in numerous nuclei of myoblasts of forming myomeres, and in their cytoplasm (inset of Figure 7C). As the myoblasts merged into myotubes in more proximal regions of the regenerating tail, a segmented immunofluorescent pattern of the elongating myomeres became evident (Figure 7D). The inter-muscle fibroblasts appeared weakly labeled or unlabeled in comparison to the elongating myotubes forming the segmented myomeres(Figures 1, 7D, E). The immunofluorescence was nuclear but was also intensely localized in the cytoplasm and in the membranes of the growing muscle fibers.In the control sections,either weak or no labelling was detected in the epidermis,ependyma, muscles, and cartilage, aside from the yellowish autofluorescence present in sparse blood vessels(Figure 7F).
DISCUSSION
General considerations
Based on transcriptome data as well as the present bioinformatics, Western blotting, and immunohistochemical results, ephrin receptor-like proteins were present in the regenerating tail of P.muralis.
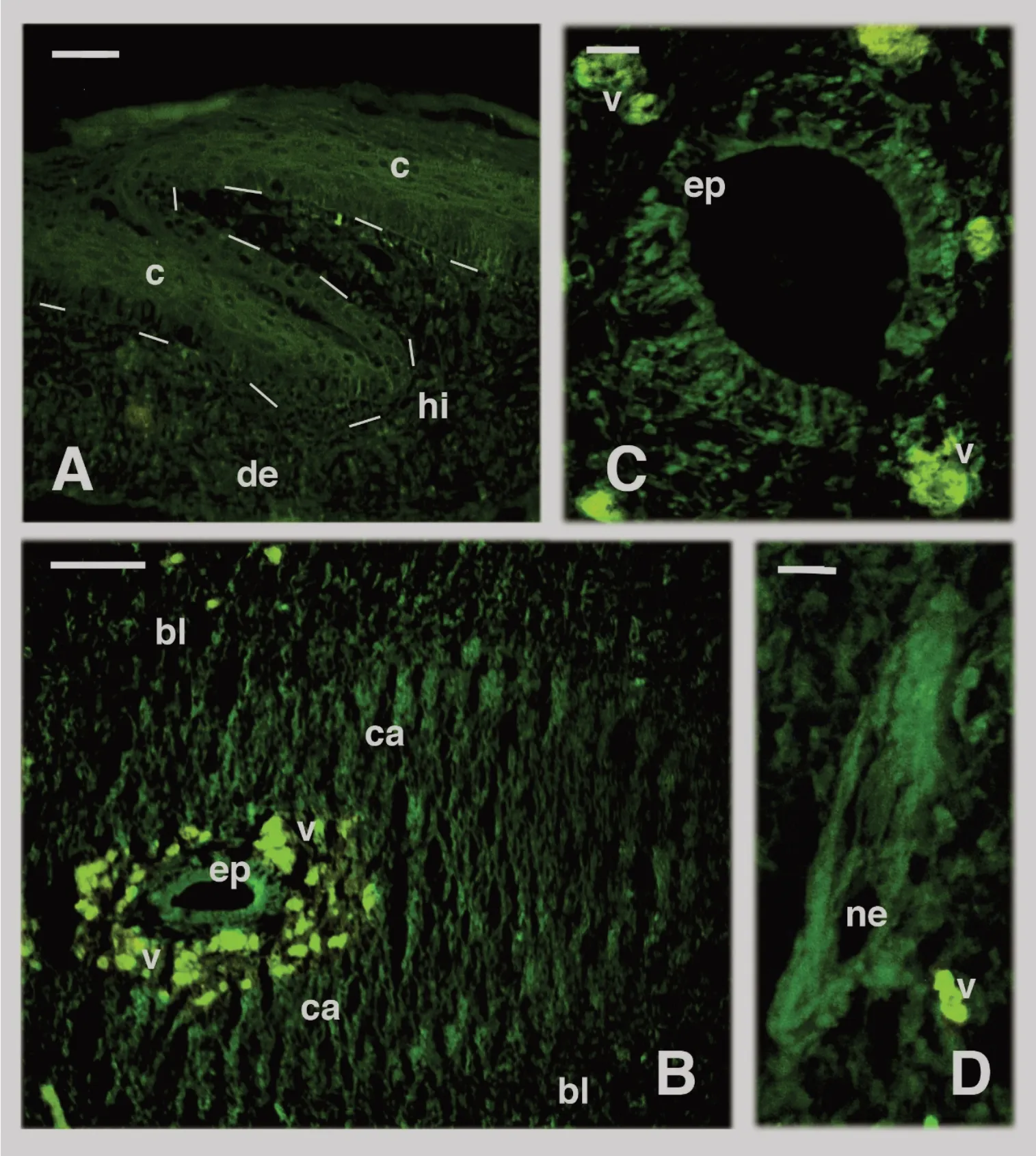
Figure 6 Immunofluorescence of tissues present in regenerating tail
Previous transcriptome study of P. muralis has indicated that numerous ephrin receptor genes are variably upregulated in the regenerating tail blastema, although their specific roles remain unknown (Vitulo et al., 2017a). They include ephb6 (ENSACAG00000008000, 8.0-fold expression),ephA4 (ENSACAG00000005061, 6.8-fold), ephb3 (ENSACAG 00000007949, 3.7-fold), ephb2 (ENSACAG00000022723, 5.8-fold), ephb1 (ENSACAG00000009241, 3.6-fold), and an unclassified ephrin type B receptor kinase gene (ENSACAG 00000008140, 3.5-fold). Among the above receptors, ephb6,ephb3, ephb2, and ephA4 are tail exclusive and are not expressed in the scarring limb (Vitulo et al., 2017a). In the regenerating lizard tail, ephrb6 has been hypothesized to function as a tumor suppressor and is considered to be a differentiating signaling protein that contrasts or regulates cell proliferation and induces differentiation in proximal areas of the regenerating blastema (Alibardi, 2017a, 2017d; Vitulo et al., 2017a). In the apical region of the blastema, numerous Wnt genes that stimulate cell proliferation are expressed,which might produce uncontrolled tumor-like proliferation without regulation. The immunolocalization of ephrin receptors in differentiating epidermal, muscle, and cartilaginous tissues supports the hypothesis of tumor-suppressor activity by ephrin receptors(Pasquale,2010).

Figure 7 Immunofluorescence of differentiating tissues in proximal areas of regenerating cones
The bioinformatics analysis of amino acid sequences between epitopes (Figure 3) suggests that the employed mouse antibody should recognize similar epitopes present in ephrin receptor-B4 (present in the database), and less so for-B3 and -B2 in A. carolinensis, and likely less also in P.muralis (although these genes are not sequenced in the latter species). The immunoblotting results showed that very low immunoreactive bands were detected within the expected molecular weight (MW) for the entire receptor, i.e., 120-140 kDa (Figure 4). Instead the main band at 60-65 kDa,partially specific as it was probably mixed with other abundant proteins of similar MW, was interpreted as a degraded form of the original protein. It remains undetermined whether this lower MW form was derived from a physiological or artifact process of degradation due to the extraction method.
The present results are partially supported by biochemical studies on nervous tissues, fibroblasts in culture, and other mouse organs, where ephrin receptor-B1 and -B2 can produce degraded forms at 65 and 45 kDa under the proteolytic action of matrix metalloproteases (Lin et al., 2012;Tomita et al., 2006). These enzymes are present in the blastema (Gilbert et al., 2015), as also shown in previous transcriptome study on P. muralis (Vitulo et al., 2017a). The protein fragments could be further processed to give rise to 15-17 kDa products.The cleavage of the intra-membrane and cytoplasmic region of the ephrin receptor operated by a γsecretase (Bong et al., 2007; Tomita et al., 2006) suggests that a cytoplasmic fragment of the receptor moves into the nucleus, although its proliferative or anti-proliferative role is not known. The nuclear localization seems to activate the transcriptional activator protein STAT3, which induces various cellular responses, including cell growth and movement, but also apoptosis(Bong et al.,2007).
In the present observation on lizard blastemas, we did not determine whether the nuclear localization seen in basal epidermal cells and muscle cells also activated specific genes and relative function. Based on the known role of complex ephrin receptors on the cytoskeleton (Klein, 2012; Palmer &Klein, 2003; Park & Lee, 2015), we suggest that possible cleavage of ephrin receptors determines intracellular responses, producing cytoskeletal rearrangements and repulsion among cells. This process induces the formation of boundaries and compartments within tissues, in particular in regenerating muscles that become segmented. Whether the 60-65 and 15-17 kDa immunoreactive bands detected in the lizard blastemas indicate biochemical processing of the ephrin receptors remains to be demonstrated as the main purpose of the Western blotting analysis was to detect the presence of immunoreactive bands in support of the immunolocalization study.
Ephrin receptors in regenerating epidermis
Ephrin-A2, -A3, and -A4 are the more common ephrin receptors found in the human epidermis, where they exert inhibitory activity on basal cell proliferation (tumor,suppressors), while stimulating keratinocyte differentiation (Lin et al., 2012; Perez-White & Getsios, 2014). These receptors likely promote cell communication and adhesion among keratinocytes, modulating the formation of cell junctions (Lin et al., 2012; Perez-White & Getsios, 2014). In the suprabasal keratinocytes of the lizard epidermis, the cytoplasmic labelling appears mainly along the perimeter of keratinocytes, reflecting the localization of this receptor. Ephrins and their receptors may be involved in the differentiation of cells that give rise to beta- and alpha-corneous layers of the scales (Alibardi, 1994,1995,2014).
During the early stages of skin wound healing in mice,between 12 h to 7 d post injury, ephrin-B1 and its receptor ephrin-B2 are detected along the cell perimeter of migrating keratinocytes that cover open wounds (Nunan et al., 2015). It is believed that this process occurs by loosening the tight and adherens junctions, but not desmosomes, facilitating the rolling movement of migrating keratinocytes along the wound border. A likely rolling movement has also been shown in the regenerating wound epidermis of lizards (Alibardi, 1994) in regions equivalent to the lateral and apical wound epidermis where ephrin-receptor immunoreactivity is present. Once the epidermis covers the tail stump, it starts to stratify and proliferate to keep pace with the expanding mesenchymal blastema that pushes elongation of the regenerating tail.
The immunolocalization of ephrin receptors, especially of ephrin receptor-B4, the subtype most likely recognized by the present antibody, in basal cell nuclei of the apical wound epidermis suggests that the receptor may be involved in the control of epidermal proliferation activity. These cells also contain most p53/63 immunolabelling, another negative regulator of cell proliferation (Alibardi, 2016). Basal keratinocytes contain high levels of proliferative markers such as telomerase, FGFreceptors, EGFreceptors, and c-myc(Alibardi, 2014, 2017a). The immunolocalization of antiproliferative (tumor-suppressors) and proliferative (oncogenes or tumor activators) proteins in the basal layer of the apical regenerative epidermis indicates that strong competition between proliferative and anti-proliferative activities is present in these cells. Colocalization of activators and suppressor proteins in the same cells has been noted in other cases,reinforcing the concept of balanced regulation (Yang et al.,2014). In the wound epidermis of the apical regions of a lizard blastema, 0.5-1.5 mm from the tip, active cell multiplication occurs along the entire epidermis, but in more proximal regions, where scales are forming, the rate of cell multiplication decreases and becomes limited to smaller regions of the forming scales (Alibardi, 1994; Wu et al., 2014).In conclusion, ephrin receptors in concert with other tumor suppressors appear essential for the control of epidermal regeneration and scale morphogenesis.
Ephrin receptors in regenerating spinal cord and nerves
Ephrins and their receptors participate in the segregation of neurons in different neuromeres of the brain and also stimulate axonogenesis in the spinal cord (Henkemeyer et al.,1994; Klein, 2012; Palmer & Klein, 2003; Park & Lee, 2015).The regenerating spinal cord in the tail of lizards includes the ependyma of the simplified spinal cord, whose cells fasciculate the descending axons derived from neurons localized in the original spinal cord and those directed toward the tip of the blastema (Alibardi & Miolo, 1995; Simpson &Duffy, 1994). Peripheral nerves are instead derived from the last three spinal sensory ganglia proximal to the regenerating blastema and from motor neurons present in the closest spinal cord segments to the blastema (Cristino et al., 2000a,2000b). Both nerves and ependymal cells contain ephrin receptors, suggesting that these proteins are also involved in axonogenesis during lizard tail regeneration.
Ephrin receptors in regenerating mesodermal tissues
The near to total absence of ephrin receptors in mesenchymal cells of the blastema indicates that these proteins are little expressed in undifferentiated cells. Conversely, the labelling of ephrin receptors in differentiating cells of forming blood vessels, myotomes, and perichondria of cartilaginous tubes suggests that these proteins are involved in the differentiation of mesodermal cells. The strong expression of ephrin receptors in segmental muscles implicates these proteins and their receptors in the determination of segmentation during regeneration, as is the case during development (Durbin et al., 1998; Stark et al., 2011). Only cells that express these receptors (myogenic) and not other types initially mixed with myoblasts, especially the fibroblasts destined to form intermuscle septa (Figure 1B, C), appear capable of merging into myotubes and myomeres. Ephrin receptors may determine the repulsion of myoblasts from fibroblasts mixed in the apical blastema, determining the initial formation of pro-muscle aggregates and later of the myomeres (Alibardi, 1995; Cox,1969; Hughes & New, 1959; Figure 1A-D). Ephrin receptors may promote the aggregation of myogenic cells that initiate to form myotubes, with the latter elongating by the incorporation of new myoblasts. The lack of expression of numerous ephrin genes in limb blastemas (Vitulo et al., 2017a) further explains the absence of regeneration and segmentation of muscle cells in this organ, which is destined to form a short scarring outgrowth.
In contrast to the muscles, no patterned distribution of ephrin receptors was observed along the differentiating cartilaginous tube and no segmentation into vertebrae occurred, although this discrepancy between muscle and cartilage morphogenesis remains unexplained. Ephrins and their receptors are involved in the formation of the initial cartilaginous condensations in developing mammalian and avian limbs (Compagni et al., 2003; Davy et al., 2004; Wada et al., 2003), and this also appeared in the lizard’s growing blastema. Condensing cartilaginous cells express ephrin-B1,whereas the forming perichondrium more intensely expresses ephrin receptor-A4. We hypothesize that an altered developmental mechanism determines the localization of ephrin-B receptors along the entire perichondrium formed around the two peripheries of the cartilaginous tube (Alibardi,2014; Lozito & Tuan, 2015, 2016; Figure 1E). This even expression of ephrin receptors is somehow correlated with the lack of segmentation of the cartilaginous tube. Future experimental studies that perturb the pattern of expression of ephrins and their receptors, especially ephrin receptors-B4 and-B6,may confirm or refute this hypothesis.
CONCLUSION
The high up-regulation of ephrin receptors detected by previous transcriptome research (Vitulo et al., 2017a) in combination with the results of the present study suggest that ephrin receptors are present in the regenerating tail. We hypothesize that these proteins act as possible tumor suppressors and differentiating factors that normalize the growth of the proximal tissues located at some distance (0.5-1.0 mm) from the tip of the tail blastema. It is likely that the ordered process leading to tail regeneration depends on crosstalk between positive genes that stimulate cell proliferation(oncogenes) and those limiting proliferation (tumor suppressors) that instead trigger differentiation. When the balance is shifted toward proliferation, as in the apical blastema, cell multiplication increases, whereas in more proximal regions anti-proliferative processes prevail and the rate of cell multiplication decreases or is limited to smaller regions of the forming scales, muscles, cartilage, nerves, and blood vessels. Ephrin receptors, in concert with other tumor suppressors (e.g., Rb, p53/63, apc), are likely essential to maintain the growth of the blastema without turning it into an uncontrolled tumor outgrowth.
COMPETING INTERESTS
The author declare that he has no competing interests.
AUTHORS’CONTRIBUTIONS
L.A.designed the study,conducted the analyses,and wrote the manuscript.
ACKNOWLEDGEMENTS
We thank Dr. F. Borsetti (Proteome Service, Department of Biology,University of Bologna) for electrophoretic analyses. We also thank the referee for help in improving the manuscript.
杂志排行

Zoological Research的其它文章
- Nest sanitation facilitates egg recognition in the common tailorbird,a plaintive cuckoo host
- A new species of Mountain Dragon(Reptilia:Agamidae:Diploderma)from the D.dymondi complex in southern Sichuan Province,China
- Postural effect on manual laterality during grooming in northern white-cheeked gibbons(Nomascus leucogenys)
- Non-invasive genetic analysis indicates low population connectivity in vulnerable Chinese gorals:concerns for segregated population management
- Genetic diversity and temporal changes of an endemic cyprinid fish species,Ancherythroculter nigrocauda,from the upper reaches of Yangtze River
- A soluble FcγR homolog inhibits IgM antibody production in ayu spleen cells