猪毛菜属3种不同光合型物种的生长适应特征比较
2019-10-25周文菲刘芙蓉姚甄业龚春梅
周文菲,刘芙蓉,姚甄业,龚春梅
(西北农林科技大学生命科学学院,陕西 杨凌712100)
干旱缺水是中国西北荒漠地区尤为突出的生态问题[1],也是影响植物生长发育和限制植物分布的重要因素[2]。植物光合作用对环境因子的变化高度敏感,因此植物受到干旱胁迫时表现出由气孔限制或非气孔限制包括光合结构发生变化、光合关键酶等活性下降所致[3]的净光合速率下降[4]。此时C3植物会通过关闭气孔的方式来保持植物体内的水分,导致细胞内CO2吸收受阻,促进核酮糖-1,5-二磷酸羧化酶/加氧酶(ribulose bisphosphate carboxylase oxygenase, Rubisco)与氧气结合,使光呼吸加强,而这一提高光呼吸或者降低CO2浓度的气孔限制会诱导C4光合的出现[5]。C4植物更适应水分胁迫和高温等逆境[6]而获得比C3植物更高的光合速率[7]。看来干旱引起的气孔和非气孔限制促进了C4光合的发生,是C4途径进化的外部动力和环境因素。随着解剖结构和酶活生理研究的深入,越来越多的证据表明随干旱加剧首先要求解剖结构发生更利于C4光合进行的变化[8-11],随后C4关键酶活性相应增强。C3植物发展出的C4途径是环境调控的产物,是对逆境的适应性进化结果,研究干旱地区植被的光合途径演化有利于对干旱地区植被的适应性机理进行深入剖析[12]。
C3、C3-C4和C4植物的生长适应性是光合途径进化的直观表现,因而对光合适应性的研究开始受到关注,朴河春等[13-14]对C3和C4植物的土壤的营养元素进行探究,陈宇琪等[15]对C3和C4植物种子的出苗和幼苗的生长等进行研究,王玉魁[16]对不同光合型物种的抗旱生理进行研究,但这些研究还不够全面系统深入。对干旱地区具有不同光合型的同属物种进行生长适应性研究非常有必要,这为完善C4光合进化理论提供了生理生态特征变化证据。猪毛菜属(Salsola)是藜科(Chenopodiaceae)中较为进化的螺胚亚科中的猪毛菜族,是该族中的原始属[17]。目前对猪毛菜属植物在种子生理、养分条件、光合指标等多个方面进行了环境适应性研究[18-21],包括对适应盐碱生境的先锋植物费尔干猪毛菜(Salsolaferganica)等的经济和生态价值研究[22]。当前国内外研究进展和本实验室的研究结果发现猪毛菜属植物中存在着不同的光合型[23-26],如蒿叶猪毛菜(S.abrotanoides)和天山猪毛菜(S.junatovii)为C3光合型,松叶猪毛菜(S.laricifolia)和白枝猪毛菜(S.arbusculiformis)为C3-C4中间光合型,木本猪毛菜(S.arbuscula)和东方猪毛菜(S.orientalis)为C4光合型,这些植物多生活在干旱荒漠及盐碱土地区,逐渐形成了适应干旱的形态特征和生理机制。因此猪毛菜属植物为研究不同光合型荒漠植物适应干旱环境的策略提供了理想材料,对促进荒漠植被恢复、改善生态环境具有重要的生态价值和社会价值。鉴于对荒漠生境不同光合型植物的抗逆生长适应特征还缺乏更全面深入的比较研究,而结合C4进化对猪毛菜属植物进行生长适应性的研究有必要进一步完善。
本研究利用DNA条形码技术构建系统进化树,选择猪毛菜属具有各自地理分布的3种不同光合型木本类猪毛菜(C3蒿叶猪毛菜、C3-C4松叶猪毛菜和C4东方猪毛菜)作为研究对象,测定其叶和种子形态结构特征、土壤和植物营养(C、N、P)、干旱响应指标[丙二醛(malondialdehyde,MDA)、可溶性糖(soluble sugar)、脯氨酸(proline)],并进行主成分分析,通过比较不同光合型植物在各自生境干旱胁迫下的生理响应,以期为更好理解C4进化提供生理生态数据支持。
1 材料与方法
1.1 试验材料
本试验以2016年采集的西北荒漠广泛分布的C3型蒿叶猪毛菜、C3-C4中间型松叶猪毛菜和C4型东方猪毛菜为研究对象,采集上述猪毛菜鲜叶样品液氮速冻带回至实验室-80 ℃保存进行生理生态相关指标测定。采样同时收集的3个物种所在的自然保护区生境概况如表1所示,随着光合型从C3经由C3-C4向C4过渡3个物种的生境年降水量依次减少,干旱程度加剧。

表1 猪毛菜属C3、C3-C4和C4光合型物种采集地的生境概况Table 1 The habitat profile of the C3, C3-C4 and C4 species of Salsola genus
1.2 试验方法
条形码基因rps16-trnQ参考Schussler等[26]的方法进行测定,采用SDS(sodium dodecyl sulfate)法提取植物DNA,参考Schussler等[26]的方法设计条形码基因rps16-trnQ的引物并进行PCR反应,送至擎科新业生物技术有限公司测序获得序列,结合NCBI(national center for biotechnology information)下载序列,并利用Mega构建NJ(neighbor-joining)进化树,采用戊二醛固定液对叶片解剖结构固定后用半薄切片机进行切片分析[26],采用体式显微镜(Nikon SMZ 25)进行拍照获得种子形态,种子重量在万分之一天平上进行测定。
采用硫代巴比妥酸法测定MDA含量[27],称取0.1 g植物样品,加入20%三氯乙酸(trichloroacetic acid,TCA)研磨冲洗并定容至2.5 mL,离心后取上清液放入离心管,再加入2 mL 0.5%硫代巴比妥酸(thiobarbituric acid,TBA),混匀后在100 ℃水浴煮10 min,离心后取上清液测定450、532和600 nm处的吸光值;采用蒽酮法测定可溶性糖含量[28],称取0.1 g植物样品,定容到7 mL水中,吸取200 μL提取液,加入800 μL蒸馏水和5 mL蒽酮试剂,在沸水浴中煮沸10 min,620 nm处测定吸光度值;采用酸性茚三酮法测定脯氨酸含量[27],称取0.1 g植物样品,用3 mL磺基水杨酸溶液,在沸水中提取10 min,吸取2 mL提取液,加入2 mL 冰醋酸和2 mL酸性茚三酮,在沸水浴中加热30 min,冷却后加入4 mL 甲苯,摇荡30 s,取上清液至10 mL离心管中,520 nm处测吸光度值。
土壤样品过0.180 mm筛子、植物叶片过0.250 mm筛子,以备用于对碳氮磷含量的分析。采用重铬酸钾外加热法测定叶片和土壤有机碳含量[29],称取0.02 g左右的叶样和0.5 g左右的土样进行测定;采用半微量凯氏定氮法测定叶片和土壤全氮含量[30],称取0.2 g左右的叶样和1 g左右的土样消煮后测定;采用钼锑抗比色法测定叶片和土壤全磷含量[31],称取0.2 g左右的叶样和0.7 g左右的土样消煮后进行测定。
1.3 数据分析
利用MEGA 7软件构建系统发育进化树,利用ImageJ2x计算叶片和种子的直径和面积等指标,土壤碳氮磷比值通过土壤养分测定的数据计算化学计量比。采用GraphPad Prism 5对数据进行分析和作图,利用SPSS 17.0和RStuido软件进行主成分分析并作图。
2 结果与分析
2.1 基于DNA条形码的猪毛菜属种系统发育树构建
DNA条形码技术是利用一段具有足够变异的标准化短基因对物种进行鉴定[32],分子系统发育进化树是通过比较生物分子序列间的关系而建立的,可以表示各个物种间的进化关系。植物DNA条形码技术有多种可供选择的基因,本研究利用NCBI(https://www.ncbi.nlm.nih.gov/)和试验获得的rps16-trnQ序列构建系统发育树,结果显示C3蒿叶猪毛菜为原始祖先,继而分别进化出C3-C4型松叶猪毛菜和C4型东方猪毛菜(图1)。为猪毛菜属不同光合型植物的进化方向和亲缘关系提供可靠证据。

图1 基于rps16-trnQ构建的猪毛菜属不同光合型物种的进化树Fig.1 Phylogenetic tree analysis based on nucleotide of rps16-trnQ
2.2 猪毛菜属3种光合型物种形态结构适应性比较
2.2.1叶片解剖结构的比较 猪毛菜属植物叶片多呈圆柱形,因而可对叶片直径进行测量从而反映叶片大小(图2a~c)。C4东方猪毛菜的叶片直径显著低于C3蒿叶猪毛菜和C3-C4松叶猪毛菜,C3-C4松叶猪毛菜的叶片直径相较于C3蒿叶猪毛菜有变小的趋势(图2d)。C3型猪毛菜叶片横切面的最外层为表皮细胞,叶肉细胞居于中间,向内是发达的贮水薄壁组织细胞,维管束分布在贮水组织中,中央的维管束较大,维管束鞘不发达;C3-C4松叶猪毛菜与C3蒿叶猪毛菜相比,其叶肉细胞的面积无显著差异,但有减少的趋势,出现具备部分功能的不完全花环结构区域,即由叶肉细胞和不连续的维管束鞘细胞组成(图2e);C4型猪毛菜具有由叶肉细胞和维管束鞘细胞组成的典型C4“花环结构”。另外,相较于C3植物,C3-C4植物和C4植物具有较大的贮水组织,且C4植物的贮水组织面积大于C3-C4植物(图2f);C3-C4植物和C4植物的栅栏组织更为发达。
2.2.2种子形态结构的变化 猪毛菜属3种光合型猪毛菜种子均为螺旋胚形(图3a~i),不同点在于种子外层覆盖物和种子大小,相较于C3植物,C3-C4植物和C4植物种子外层覆盖物更为紧密;C3-C4植物的种子大小和重量都明显大于C3植物和C4植物(图3j,k)。图3a~c分别显示C3型蒿叶猪毛菜的上下两面和剥去覆盖物后的种子形态;图3d~f分别显示C3-C4植物松叶猪毛菜的上下两面和剥去覆盖物后的种子形态;图3g~i分别显示C4植物东方猪毛菜的上下两面以及剥去覆盖物后的种子形态。
2.3 猪毛菜属3种光合型物种生理水平适应性比较
C4型东方猪毛菜的丙二醛含量略高于C3-C4型松叶猪毛菜,但都明显低于C3型蒿叶猪毛菜,说明C3-C4型松叶猪毛菜和C4型东方猪毛菜在干旱环境下的膜稳定性都较高(图4a);C3-C4型松叶猪毛菜的可溶性糖含量低于C3蒿叶猪毛菜和C4东方猪毛菜(图4b);随着光合型向C4转变,脯氨酸含量呈现逐渐下降的趋势(图4c)。推测可溶性糖在C4东方猪毛菜的渗透调节中起重要作用,脯氨酸在C3-C4松叶猪毛菜渗透调节中起一定作用,而可溶性糖和脯氨酸在C3型蒿叶猪毛菜通过渗透调节的抗旱适应性中均起一定作用。
2.4 猪毛菜属3种光合型物种生境贫瘠适应性比较
2.4.1植物叶生态化学计量特征 C3-C4松叶猪毛菜的碳含量低于C3蒿叶猪毛菜,但二者都远远低于C4东方猪毛菜(图5a);C3植物的氮含量显著高于C3-C4植物,但是两者都显著低于C4植物(图5b),这与磷含量在3种光合型猪毛菜中的趋势类似(图5c)。C3蒿叶猪毛菜和C4东方猪毛菜的C/N和C/P无显著差异,但是都显著低于C3-C4松叶猪毛菜(图5d,e);这说明C3-C4松叶猪毛菜的N、P元素利用率最高,即同化单位N或者P元素固定的碳含量最高。再结合3种光合型植物的碳含量发现不同光合型猪毛菜采用不同策略来增强固碳量,C3-C4松叶猪毛菜采用的是增强N、P元素的利用率来增加植物的固碳量,而C4东方猪毛菜通过增加N、P元素的含量来增加植物的固碳量。从C3到C3-C4再到C4猪毛菜,叶N/P呈现下降趋势(图5f),表明这3种光合型猪毛菜体内P起限制作用,到C4东方猪毛菜,这种限制作用逐渐减小,暗示C4进化过程中低磷胁迫越来越弱。同时N/P下降也反映出植物可以降低对氮的需求,以便更能适应贫瘠的环境从而提高耐瘠薄性。

图2 猪毛菜属3种不同光合型物种的叶片解剖结构特征比较Fig.2 The leaf anatomical structure and relative quantitative index of Salsola genus 不同小写字母表示差异显著(P<0.05)。下同。Different small letters indicate significant differences (P<0.05). The same below.

图3 猪毛菜属不同光合型物种的种子形态差异以及直径和百粒重的比较Fig.3 The variation of seeds morphology, 100-seed weight and diameter among 3 photosynthetic types of Salsola genus
2.4.2土壤生态化学计量特征 C3蒿叶猪毛菜生境中的土壤碳、氮含量显著高于C3-C4松叶猪毛菜和C4东方猪毛菜,C3-C4松叶猪毛菜和C4东方猪毛菜之间差异不大(图6a,b),说明C3-C4松叶猪毛菜和C4东方猪毛菜生境中土壤的营养元素相对匮乏。C3-C4松叶猪毛菜生境中的土壤磷含量显著高于C3蒿叶猪毛菜和C4东方猪毛菜,C3蒿叶猪毛菜和C4东方猪毛菜之间差异不大(图6c)。C3-C4松叶猪毛菜的土壤C/N略高于C3蒿叶猪毛菜和C4东方猪毛菜,但不显著,C3蒿叶猪毛菜和C4东方猪毛菜之间的C/N差异不大(图6d),有效氮含量略低于全国平均水平。C4东方猪毛菜的土壤C/P和土壤N/P略高于C3-C4松叶猪毛菜,但是它们的土壤C/P和土壤N/P都显著低于C3蒿叶猪毛菜(图6e,f),猪毛菜生境土壤C/P含量明显低于全国平均水平,且C3-C4松叶猪毛菜和C4东方猪毛菜的土壤有效磷含量远远大于C3蒿叶猪毛菜。

图4 猪毛菜属不同光合型物种的抗旱性差异比较分析Fig.4 The physiological index among the 3 photosynthetic types of Salsola genus

图5 猪毛菜属不同光合型物种叶营养元素及比值变化Fig.5 The content of leaf nutrient elements and its ratio of Salsola genus
2.5 主成分分析
逆境下植物的生长适应性由多种生理生态指标共同塑造,而这些指标之间具有一定的相关性,为探究这些相关指标对猪毛菜适旱生长的贡献率,本试验采用主成分分析法,对干旱驱动的不同光合型猪毛菜的各项指标进行综合分析,为C4进化的表观生长影响因子进行归类并评价。
通过主成分分析,提取2个主成分,在第一主成分轴上,C3-C4植物松叶猪毛菜和C4植物东方猪毛菜得分较高。在第二主成分轴上,C3植物蒿叶猪毛菜得分较高(图7)。第一主成分的方差贡献率为51.079%,在这一主成分中,叶片C、N、P含量和叶片C/N、C/P、种子直径和百粒重、可溶性糖含量和土壤C/N、土壤P含量等指标具有较高的载荷值,对松叶猪毛菜贡献较大(表2和图7);第二主成分的方差贡献率为48.921%,这一主成分中,叶片C、N和P以及贮水组织面积、维管束鞘面积、可溶性糖含量、降水量等具有较高的载荷值,对东方猪毛菜贡献较大(表2)。
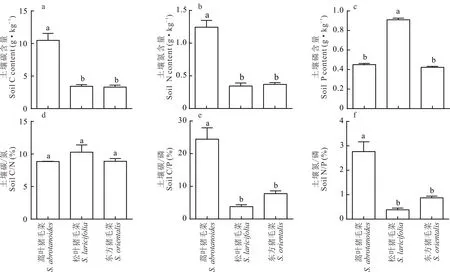
图6 猪毛菜属不同光合型物种生境土壤营养元素及比值变化Fig.6 The content of soil nutrient elements and its ratio among 3 photosynthetic types of Salsola genus
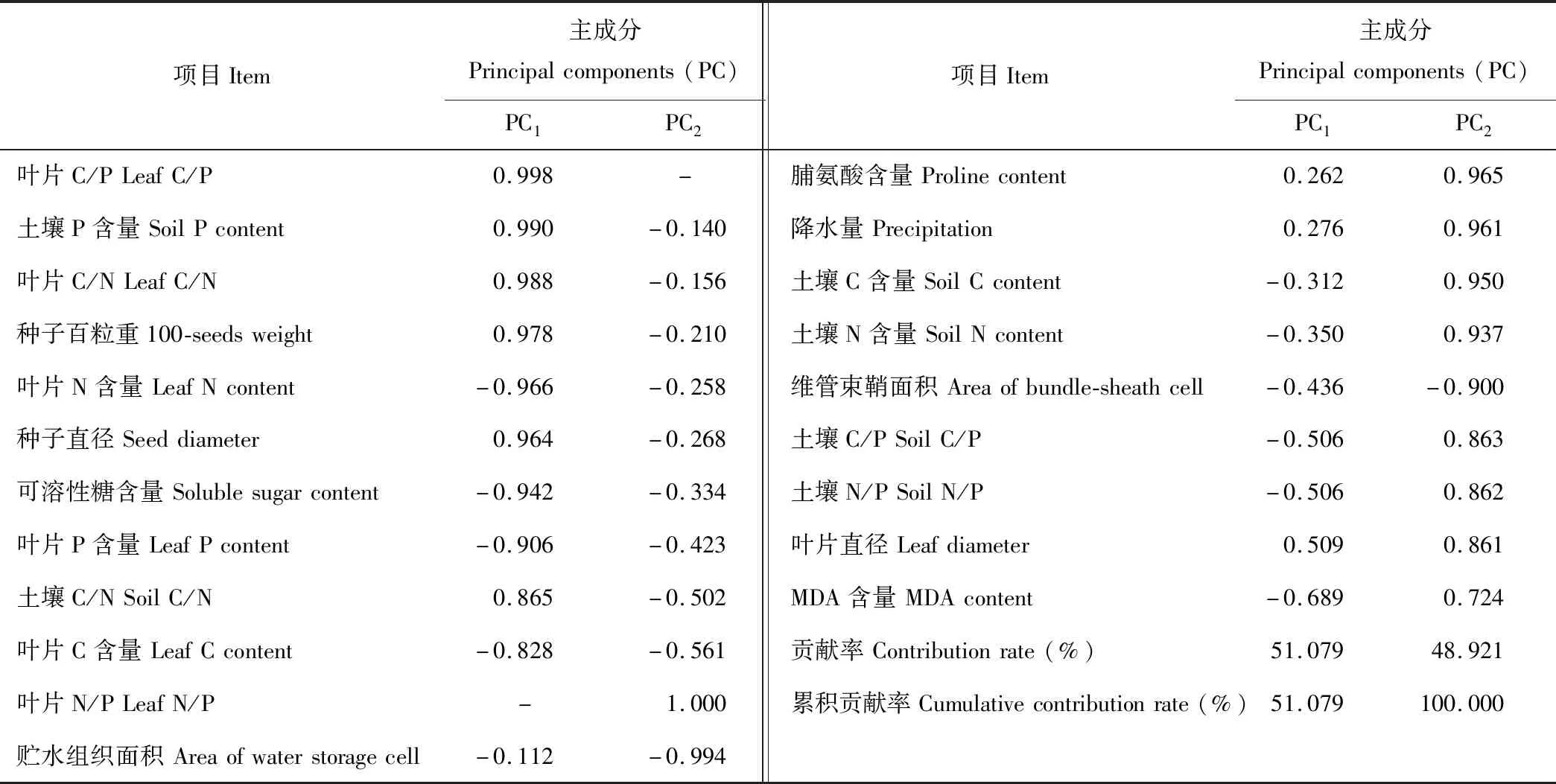
项目Item主成分Principalcomponents(PC)PC1PC2项目Item主成分Principalcomponents(PC)PC1PC2叶片C/PLeafC/P0.998-脯氨酸含量Prolinecontent0.2620.965土壤P含量SoilPcontent0.990-0.140降水量Precipitation0.2760.961叶片C/NLeafC/N0.988-0.156土壤C含量SoilCcontent-0.3120.950种子百粒重100-seedsweight0.978-0.210土壤N含量SoilNcontent-0.3500.937叶片N含量LeafNcontent-0.966-0.258维管束鞘面积Areaofbundle-sheathcell-0.436-0.900种子直径Seeddiameter0.964-0.268土壤C/PSoilC/P-0.5060.863可溶性糖含量Solublesugarcontent-0.942-0.334土壤N/PSoilN/P-0.5060.862叶片P含量LeafPcontent-0.906-0.423叶片直径Leafdiameter0.5090.861土壤C/NSoilC/N0.865-0.502MDA含量MDAcontent-0.6890.724叶片C含量LeafCcontent-0.828-0.561贡献率Contributionrate(%)51.07948.921叶片N/PLeafN/P-1.000累积贡献率Cumulativecontributionrate(%)51.079100.000贮水组织面积Areaofwaterstoragecell-0.112-0.994
从载荷图(图7)可知,在第一主成分上能够分开C3蒿叶猪毛菜和C3-C4松叶猪毛菜,在第二主成分上能够分开C3蒿叶猪毛菜和C4东方猪毛菜。C3蒿叶猪毛菜的土壤C、N含量和土壤N/P、C/P、丙二醛和脯氨酸含量较高;C3-C4松叶猪毛菜的种子直径和百粒重、叶片C/N、C/P较高;C4东方猪毛菜的维管束鞘面积和贮水组织面积、叶片C、N、P含量、可溶性糖含量较高。
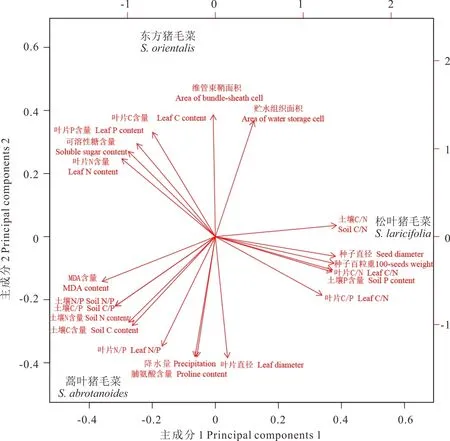
图7 猪毛菜属不同光合型物种多种指标的载荷图Fig.7 Load map of various indexes among 3 photosynthetic types Salsola genus
3 讨论
至此,已对藜科猪毛菜属3种不同光合型植物应对干旱贫瘠的生长适应性做了初步比较。
首先确立不同光合型猪毛菜的亲缘关系和C4演化方向。已有研究对菊科黄顶菊属(Flaveria)不同光合型植物的关系进行研究,发现黄顶菊属C3植物位于系统发育树的最底部,之后进化出C3-C4和C4光合途径[33]。本研究利用DNA条形码对不同光合型猪毛菜植物进行构树聚类研究,发现C3-C4型猪毛菜与C4型猪毛菜也是由C3型猪毛菜演化而来,这与不同光合型黄顶菊属植物的进化总体方向一致[34],明确了不同光合型猪毛菜的亲缘关系和C4进化方向。
不同光合型猪毛菜应对干旱胁迫产生叶和种子的形态结构不同。叶片可以进行光合作用和蒸腾作用等多种生命活动,在植物生长过程中发挥重要作用,在行使生理功能的过程中叶片与环境之间有着紧密的关系,因而为了适应环境的变化,植物叶片会在外部形态、解剖结构等多个方面发生一系列的适应性变化。猪毛菜属3个种从C3向C4方向过渡,外部形态上柱状叶趋于细短,细小的叶型可以减少水分蒸发面积而更耐旱[4],表明猪毛菜属植物在向C4进化过程中其叶片外部形态越来越适应干旱环境。解剖结构上相较于C3种,C3-C4种和C4种的栅栏组织和贮水组织更为发达。栅栏组织/海绵组织越大,即栅栏组织越发达,植物越耐旱;贮水组织越发达,植物越耐旱[35-36]。这便是猪毛菜C4植物更耐旱的原因[37]。同时猪毛菜C4种叶片具有典型的猪毛菜型“花环”结构,即由外层整齐排列的叶肉细胞和内层较发达的维管束鞘细胞组成,旨在通过四碳化合物从叶肉细胞向维管束鞘的运输实现二氧化碳在维管束鞘细胞内的再固定,为高光效光合作用提供了快速产生和运输有机产物的物质条件和合理结构。
干旱条件下植物的生长环境非常严酷,因此,为适应这种严酷的条件,植物具有其特殊的适应机制,其中种子繁殖便是荒漠植物产生的一种适应机制[38]。从种子自身萌发特性而言,种子大小影响植物生长和存活:较大的种子萌发缓慢但萌发后生存力强,自身竞争力强;较小种子萌发较快但萌发后生存力较弱,但种子大小与种子数量和种子扩散分布成反比,具有较小种子的植物,其种子在数量上一般居多,采取的是机会主义生存策略[39]。C3-C4松叶猪毛菜的种子无论在形态上还是重量上都明显大于C3蒿叶猪毛菜和C4东方猪毛菜,其采用增强种子自身竞争力的机制来适应环境,而C3蒿叶猪毛菜和C4东方猪毛菜则采取在种子传播上占据优势的策略。为了适应干旱的环境,荒漠植物种子表面具有水溶性抑制物质,只有在大量降水后,这些抑制物质被洗脱掉才能萌发,以保证形成的幼苗不致因缺水而枯死。从形态上可以看出来,猪毛菜种子外有一层覆盖物,这层覆盖物可能发挥着保护种子免受环境胁迫、保持种子水分等作用,随着向C4进化,这层覆盖物越加致密发达。
干旱胁迫下植物细胞膜容易发生过氧化反应产生丙二醛,因此常用叶片的丙二醛含量来反映植物膜脂过氧化程度,即丙二醛含量越高,说明细胞膜不稳定,则植物的抗旱能力越弱。不同光合型猪毛菜应对干旱胁迫在抗性生理方面表现出差异。干旱胁迫下植物叶片容易遭受氧化胁迫使得膜脂过氧化产生丙二醛,但耐旱植物在逆境中产生的丙二醛含量较低[40],本试验中C4物种产生的丙二醛含量较低。干旱胁迫下,植物细胞内水势较低,会通过积累渗透调节物质来吸收更多的水分以维持植物体正常的生命活动,这些渗透调节物质主要有可溶性糖和脯氨酸等。本研究结果中,脯氨酸和可溶性糖在C3蒿叶猪毛菜中的作用均大于C3-C4蒿叶猪毛菜,相较于脯氨酸,可溶性糖在C4东方猪毛菜中发挥重要的作用。不同植物在干旱胁迫下积累的渗透调节物质种类存在差异,如无机离子在中生植物渗透调节中起的作用较小[41],但在葡萄(Vitisvinifer)植株中却起着大于可溶性有机物质的作用[42]。可溶性糖含量在甜菜(Betavulgaris)[43]、豌豆(Pisumsativum)[44]等植物抗旱中发挥着重要的渗透调节作用,但是也有研究指出干旱胁迫下,植物碳同化率降低,可溶性糖含量降低[45],本试验中相较于脯氨酸,可溶性糖在调节C4东方猪毛菜渗透作用的过程中起着更为重要的作用,在向C4光合过渡中植物的光合能力逐渐增强,通过可溶性糖来调节植物体内的渗透势具有极为便利的物质基础。
植物通过光合作用吸收大气中的二氧化碳,通过叶的蒸腾作用获取土壤中的水分和矿质营养元素,驱动陆地生态系统中水和碳氮磷等元素的生物地化循环[46]。作为植物的基本营养元素,C元素是构成植物体最主要的结构元素,而N和P是植物体主要的功能元素,因为它们是植物体内各种蛋白质和遗传物质重要的组成元素,C、N、P在植物生长和各种生理调节机能中关系密切,且发挥着重要作用。植物叶片中的C/N、C/P不仅可以反映植物N、P元素的利用率,还反映了植物吸收单元同化C元素的能力。换言之,比值越高,说明植物吸收单位N或者P元素同化的C元素越多[47-48],植物N/P则被用来评价生态系统中营养元素的限制作用。本试验结果中,C3-C4植物的C/N、C/P中明显大于C3植物和C4植物,这说明C3-C4植物的N、P元素利用率最高,即同化单位N或者P元素固定的C含量最高,再结合3种光合型植物的碳含量发现不同光合型猪毛菜采用不同策略来增强碳积累,C3-C4松叶猪毛菜采用的是增强N、P元素的利用率来增加植物的碳积累,而C4东方猪毛菜通过增加N、P元素的含量来增加植物的碳积累。植物组织中的营养元素含量被用来评价生态系统中营养元素限制作用,N/P的化学计量比值被用来评价N或P中哪一种更多地起限制生物量产量和物种丰度的作用,一般认为,N起限制作用的N/P临界值为6.7~16.0,而P起限制作用的N/P临界值为12.5~26.3,低于临界值表示N起限制作用,高于临界值表示P起限制作用[49]。本试验结果表明,这3种光合型猪毛菜体内N/P所处范围属于P起限制作用,且随着向C4进化,这种限制作用逐渐减小,有研究表明低磷胁迫能显著降低叶片相对含水量和叶绿素含量,同时,低磷胁迫能显著降低叶片光合速率和气孔导度[14],因而C4进化过程中低磷胁迫越来越弱。
不同光合型猪毛菜应对贫瘠生境在生态化学计量特征方面表现出差异。结合所在区域生境土壤的营养元素,能揭示植物对营养元素的需要和当地土壤的养分供给能力,对植物与环境的适应与反馈能力都具有十分重要的意义[50]。土壤是植物生存的物质基础和重要的环境条件,土壤C、N、P等养分含量是衡量土壤肥沃程度的重要指标,本试验中,相较于C3蒿叶猪毛菜,C3-C4松叶猪毛菜和C4东方猪毛菜生境中土壤的营养元素相对匮乏。同时,土壤C/N、C/P和N/P也是确定土壤C、N、P平衡特征的重要参数,全球和全国土壤C/N平均水平为14.3和10~12[51],C3-C4松叶猪毛菜的C/N大于C3蒿叶猪毛菜和C4东方猪毛菜,但都维持在全国平均水平附近,土壤有机层的C/N较低表明土壤有机层的有效氮含量相对较高,因此尽管C3-C4松叶猪毛菜和C4东方猪毛菜生境土壤C、N含量较低,但土壤有效氮含量与C3植物相当;陆生生态系统中,生物固氮量随着土壤中N/P增大而减少,随土壤中N/P减少而增大[52],本试验中C3-C4松叶猪毛菜和C4东方猪毛菜其生境土壤的生物固氮量明显大于C3植物;较低的C/P也反映了土壤中有效磷浓度较高,我国土壤C/P的平均值为61[53],猪毛菜生境中C/P明显低于全国平均水平,从叶片营养元素分析中可看出猪毛菜植物受到低磷胁迫,C3-C4较低的土壤C/P也反映了土壤中有效磷浓度较高,松叶猪毛菜和C4东方猪毛菜的土壤有效磷浓度远远大于C3植物。这一结论与猪毛菜叶片存在低磷胁迫且C3-C4松叶猪毛菜和C4东方猪毛菜能缓解低磷胁迫相一致。简而言之,C3-C4物种松叶猪毛菜生境中土壤C、N含量贫瘠,但土壤P含量丰富且土壤有效P含量较高,叶片C、N、P含量较低,但其叶片P元素利用率较高;C4植物东方猪毛菜生境中土壤C、N含量贫瘠,但土壤有效P含量较高,叶片C、N、P含量较高。表明C3-C4植物松叶猪毛菜和C4植物东方猪毛菜采用不同的策略来缓解低磷胁迫。
C4进化的驱动因素之一是干旱,其形成受环境因子以及植物对环境的生物和生态适应性差异的影响,通过对这些指标进行主成分分析发现,在C4进化中,主要受两大类因素影响,各因素的方差贡献率分别为51.079%和48.921%。在第一主成分上能够分开C3蒿叶猪毛菜和C3-C4松叶猪毛菜,在第二主成分上能够分开C3蒿叶猪毛菜和C4东方猪毛菜。C3植物在第二主成分上具有较高的得分,表明相较于C3植物,C3-C4植物和C4植物更能适应贫瘠的环境。
4 结论
本试验基于 DNA 条形码对 3 种光合型猪毛菜的生长适应性包括形态结构、抗旱生理和生态特征进行比较和评价,获得以下主要结论,为完善C4进化理论提供表观生长适应证据。
1)C3光合型猪毛菜是原始祖先,C3-C4中间型猪毛菜与C4光合型猪毛菜由C3光合型猪毛菜进化而来;2)与C3光合型蒿叶猪毛菜相比, C3-C4中间型松叶猪毛菜以及C4光合型东方猪毛菜叶从外部形态和解剖结构上逐渐形成适应干旱的结构特征,且种子的传播策略也因种子大小不同而发生变化;3)C4光合型猪毛菜因高光合能力使其具备通过可溶性糖来调节渗透能力的物质基础,从而提高抗旱能力;4)C3-C4型猪毛菜是通过提高氮和磷的利用效率来增加固碳量,C4光合型猪毛菜是通过氮和磷含量的增多来增加固碳量。相较于C3物种蒿叶猪毛菜,C3-C4物种松叶猪毛菜和C4物种东方猪毛菜更能适应贫瘠的环境。
