2014—2018 年猪流行性腹泻病毒N 基因遗传变化分析
2019-10-25邢广旭焦文强刘运超王治方王克领徐引弟李海利张改平
邢广旭,焦文强,刘运超,王治方,王克领,徐引弟,李海利,张改平
(1.河南省农业科学院动物免疫学重点实验室,河南郑州450002;2.河南省农业科学院畜牧兽医研究所,河南郑州450002;3.河南省畜禽繁育与营养调控重点实验室,河南郑州450002)
猪流行性腹泻(Porcine epidemic diarrhea,PED)是由猪流行性腹泻病毒(Porcine epidemic diarrhea virus,PEDV)感染猪引起的一种高度接触性肠道传染性疾病,临床主要表现为呕吐、水样腹泻,最后脱水死亡。该病可感染各年龄阶段的猪,7 日龄尤其是3 日龄以内的哺乳仔猪发病后死亡率最高可达100%[1]。该病在1971 年由英国首次报道[2],并且迅速传播到欧洲其他国家,并在比利时流行期间被证实其病原为PEDV,给欧洲的养猪业带来了重大损失[3]。亚洲的日本、韩国、泰国等国家都曾先后报道过PED 的流行[4-8]。我国学者宣华等[9]于1984 年首次报道PEDV 在我国猪场的存在,并且在2010 年以前主要是以散发为主。但是从2010 年10 月开始,我国南部多省份暴发了仔猪严重腹泻,并迅速蔓延到全国多个省份和地区,测序结果证明,PED暴发是由变异的PEDV 引起[10-12];2013 年5 月,美国暴发了PED 疫情,并很快蔓延到美国全国,其分离株与我国安徽分离株(AH20122)亲缘关系最为接近,表明PEDV 突变重组现象非常严重,同时这也是现有疫苗难以提供免疫保护的主要原因[13]。
PEDV 属于冠状病毒科(Coronaviridae)冠状病毒属(Coronavirus),核酸类型为单股正链RNA[14]。基因 组 由5′非 编码 区(untranslated region,UTR)、3'UTR 以及7 个开放阅读框(open reading frames,ORFs),分别编码3 个非结构蛋白(non-structural proteins,NSPs)和4 个结构蛋白(structural proteins,SPs)组成,也就是S 蛋白、E 蛋白、M 蛋白和N 蛋白[15]。其中,N 蛋白是所有结构蛋白质中含量最多的蛋白,在PEDV 感染早期机体会产生大量针对N 蛋白的抗体,因此,N 蛋白被用于早期诊断[16]。N 蛋白在宿主细胞内诱导产生细胞免疫过程中扮演着非常重要的角色[17]。此外,N 基因参与病毒基因组的转录、病毒核衣壳的形成以及病毒RNA 的包装[18-19]。
为深入了解PEDV 在我国遗传变异的情况,本研究对2014 年10 月至2018 年3 月采集到的205 份PEDV 疑似病料进行RT-PCR 检测,扩增阳性样品的N 基因序列并进行测序,旨在为指导临床PEDV的防控工作提供参考。
1 材料和方法
1.1 病料采集
本研究于2014 年10 月至2018 年3 月共采集来自河南、河北、山东、山西等地区疑似PEDV 病料205 份,包括仔猪小肠组织、粪便以及母猪血液等。
1.2 主要试剂
Taq DNA聚合酶、鼠源反转录酶(M-MLV)、RNA酶抑制剂、pMD18-T Simple 载体、DL2000 Marker RNA 提取试剂盒,均购自大连宝生物工程有限公司;琼脂糖购自Oxoid 公司;其他试剂均为国产分析纯。
1.3 引物设计
参考GenBank 中收录的PEDV 毒株N 基因序列,使用Primer Premier 5.0 软件设计引物,上游引物为:5′-TACGGATCCATGGCTTCTGTCAGC-3′,下游引物为:5′-TCGCTCGAGTTAATTTCCTGTATC-3′,引物由生工生物工程(上海)股份有限公司合成。
1.4 PEDV 的N 基因的克隆与测序
按照宝生物RNA 提取试剂盒操作说明书完成疑似样品的RNA 提取,随即合成cDNA,反应体系为:5×NLV 反转录缓冲液2 μL,dNTP 2 μL,MLV 反转录酶0.5 μL,rNAse 抑制剂0.3 μL,总RNA4.9 μL,充分混匀后置离心机瞬时离心混匀,42 ℃1.5 h,70 ℃10 min。以cDNA 为模板进行PCR 扩增,PCR反应体系为:Taq DNA 聚合酶1 μL,cDNA 1 μL,上下 游 引 物 各1 μL,dNTP 4 μL,10×PCR Buffer(Mg2+plus)5 μL,无菌双蒸水补足50 μL。反应程序为:94 ℃预变性5 min;94 ℃30 s,57 ℃30 s,72 ℃1 min,35 个循环;最后72 ℃延伸10 min。扩增结束后取5 μL PCR 产物于1.0%琼脂糖凝胶电泳检测扩增结果,回收与预期大小一致的PCR 产物,并与pMD18-T simple 载体连接,4 ℃过夜后转化至JM109 感受态细胞,蓝白斑筛选后挑取单克隆进行培养,并提取质粒送至生工生物工程(上海)股份有限公司进行测序。
1.5 数据处理
使用MEGA 5.0 软件构建系统进化树。
2 结果与分析
2.1 PEDV 的N 基因的扩增

205 份待检样品中,有191 份样品检测为PEDV 阳性,阳性率高达93.17%。从图1 可以看出,PCR 扩增PEDV N 基因的引物可以扩增出1 326 bp大小的片段,片段符合预期大小。
2.2 PEDV 的N 基因比对结果
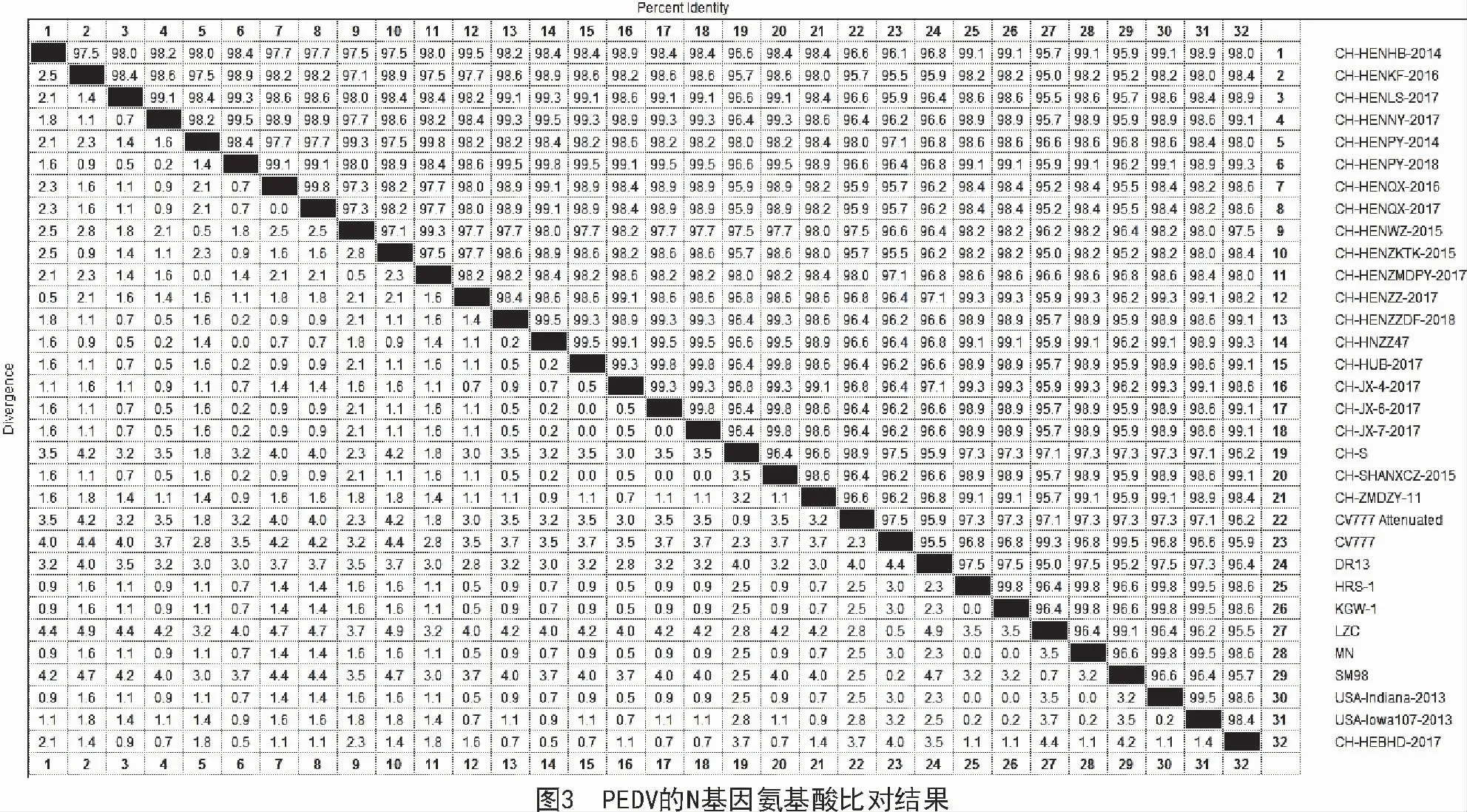
本研究得到的19 株N 基因序列与国内外参考毒株进行比对,发现这19 株N 基因序列与国内外代表毒株相比,其核苷酸的同源性为94.8%~99.5%(图2),氨基酸的同源性为95.0%~99.3%(图3)。

2.3 PEDV 的N 基因的进化分析
使用MEGA 5.0 软件,将本研究得到的19 株PEDV 的N 基因序列与GenBank 中下载的序列构建系统进化树(图4)。从图4 可以看出,本研究得到的19 株PEDV 的N 基因序列与CV777、LZC、SM98、CH-S 和DR13 等毒株不在同一个分支上,表明这19 株N 基因序列与传统经典毒株进化关系较远;而CH-HENWZ-2015、CH-HENPY-2014、CH-HENZMDPY-2017 3 株序列形成一个分支,为G2b 亚群;其余16 株N 基因序列形成另外一个分支,为G2a亚群。表明N 基因进化趋势日益复杂,需要加强此方面的监控。

3 结论与讨论
通过RT-PCR 方法,本研究得到了19 株PEDV 的N 基因序列,与先前的报道一致。本研究得到的PEDV 的N 基因由1 326 个核苷酸组成,共编码441 个氨基酸,分子量为56 ku 左右,没有任何碱基的缺失和插入。有研究显示,PEDV 基因组有4 个超变区[20],其中,V4 位于N 蛋白内,这与以前的结论不一致[21]。
本研究得到的19 株PEDV 的N 基因序列与GenBank 中下载的参考序列核苷酸的同源性为94.8%~99.5%,氨基酸的同源性为95.0%~99.3%。考虑到本研究所采集的样品来自河南、河北、山西、山东、湖北、甘肃、江西、上海等多个省市,并且本研究所采集的样品一共205 份,样本数量足够大,而且检测出191 份PEDV 阳性样品,所以,不管是样品的采集数量还是样品的覆盖面积,均具有较强的说服力,证明N 基因是PEDV 基因组中很保守的一个结构蛋白。氨基酸比对结果发现,本研究得到的19 株N 蛋白氨基酸序列主要发现以下位点的突变:K123-N123,A142-T142,R241-K241,K252-R252,N255-S255,M394-T394,E400-D400,A408-L408,V402-S402,从而从氨基酸水平证实N 基因缺失很保守。
采用MEGA 5.0 软件构建的基于N 基因的系统进化树发现本研究得到的19 株N 基因序列与CV777、LZC、SM98、CH-S 和DR13 等国内外代表性毒株不在同一个分支上,表明这19 株N 基因序列与传统经典毒株进化关系较远;其中,16 株N 基因序列与MN、USA-Indiana-2013、USA-Iowa107-2013、HRS-1、KGW-1、CH-ZMDZY-11 形成一个分支,为G2a,CH-HENWZ-2015、CH-HENPY-2014、CHHENZ-MDPY-2017 这3 株序列形成另一个分支,为G2b。综上所述,表明我国目前PEDV 的N 基因与以往流行的经典毒株亲缘关系较远,大部分毒株与目前国内外分离的毒株在一个进化分支中,部分毒株独立进化成一个分支。表明PEDV 遗传进化趋势日益复杂,需要长期监控。
