混种少花龙葵嫁接后代对镉胁迫枇杷幼苗光合生理的影响①
2019-10-19李红艳陈发波黄佳璟廖明安林立金李华雄
李红艳,陈发波,黄佳璟,温 铿,廖明安,林立金*,蒋 伟,李华雄
混种少花龙葵嫁接后代对镉胁迫枇杷幼苗光合生理的影响①
李红艳1,陈发波2,黄佳璟3,温 铿3,廖明安3,林立金1*,蒋 伟4,李华雄5
(1 四川农业大学果蔬研究所,成都 611130;2 长江师范学院生命科学与技术学院,重庆 408100;3 四川农业大学园艺学院,成都 611130;4 成都师范学院化学与生命科学学院,成都 611130;5 四川省内江市农业科学院林果研究所,四川内江 641000)
采用盆栽试验,将4种少花龙葵嫁接后代分别和枇杷(大五星枇杷和川早枇杷)幼苗混种于镉含量为10 mg/kg的污染土壤中,研究了混种对两种植物光合生理的影响。结果表明:混种后,枇杷幼苗相应的叶绿素a含量、叶绿素b含量、叶绿素总量、净光合速率、蒸腾速率、胞间CO2浓度、气孔导度及可溶性糖含量均高于单种,叶表面蒸汽压亏缺降低;少花龙葵嫁接后代相应的SPAD值和净光合速率、蒸腾速率、胞间CO2浓度、气孔导度均高于单种,其可溶性糖含量较单种有所降低,混种大五星枇杷的少花龙葵嫁接后代的叶表面蒸汽压亏缺高于其单种,但混种少花龙葵嫁接后代的川早枇杷叶表面蒸汽压亏缺低于其单种。因此,少花龙葵嫁接后代混种枇杷可提高两种植物的光合作用,进而可促进两种植物的生长。
少花龙葵嫁接后代;枇杷;光合生理;混种;镉
近年来,由于工业“三废”的排放以及农药化肥的过度使用,果园土壤镉污染越来越严重[1-3]。在镉胁迫下,植物细胞内的糖、脂、蛋白质等生物大分子会发生一系列变化,对植物有一定毒害作用[4]。另外,Cd2+会通过影响植物的气孔导度、叶绿素含量以及光合作用器官,进而抑制植物的光合作用,降低经济作物的品质与产量[5]。混种是一种具有悠久历史的农艺措施,能够充分利用各种自然资源,实现光照、水、养分的集约化利用[6-7]。在玉米、小麦等农作物的种植过程中利用不同混种模式是提高粮食产量的重要栽培手段[8-9]。超富集植物对重金属具有极强的耐性,对重金属的吸收量超过普通植物 100 倍以上,常被应用于重金属污染土壤的修复中[10-11]。已有研究发现,在重金属胁迫下混种超富集植物能够有效提高经济作物的生物量及叶绿素含量,同时对土壤脲酶、蔗糖酶、过氧化氢酶的活性也有一定促进作用[12-13]。
枇杷((Thunb.) Lindl.)为蔷薇科,枇杷属植物,其花、果、叶、根均可入药,是一种药食兼用的宝贵资源[14]。枇杷是一种浅根系植物,80% 的吸收根系分布在10 ~ 50 cm 的土层中[14]。有关研究表明,镉胁迫会造成枇杷光合效率降低,净光合作用下降,进而影响枇杷的生长发育[15]。少花龙葵是一种潜在的镉超富集植物,其根系发达,主要分布在0 ~ 30 cm 的土层中,最深可达1 m土层[16]。将少花龙葵和枇杷混种,两种植物在根系分布上相互重叠,能够充分发挥“根际对话”的作用[17]。石军等[18]研究表明,在土壤镉含量约为10 mg/kg 时,大五星枇杷和川早枇杷幼苗的生长受到抑制,其生物量均显著降低,说明10 mg/kg的土壤镉对枇杷幼苗造成了比较严重的胁迫作用。因此,本研究以少花龙葵嫁接后代和枇杷幼苗为材料,将两种植物混种于10 mg/kg的镉污染土壤中,研究混种后少花龙葵嫁接后代和枇杷幼苗的光合生理特性,以期筛选出在镉胁迫土壤条件下能够有效提高枇杷幼苗光合作用的少花龙葵嫁接后代,为促进枇杷幼苗的生长发育提供理论依据。
1 材料与方法
1.1 试验材料
2013年5月,将采自四川省雅安市汉源县唐家乡小关村(29°24′ N,102°38′ E)枇杷园的两年生大五星枇杷实生幼苗和川早枇杷实生幼苗假植于四川农业大学雅安校区农场(29°59′ N,102°59′ E)果园中。小关村平均海拔约800 m,地处北温带与季风带之间的山地亚热带气候区,具有典型的干热河谷气候特点,多年平均气温17.9 ℃,多年平均降雨量741.8 mm,多年平均日照1 475.8 h,多年平均蒸发量1 248.2 mm。四川农业大学雅安校区农场位于四川省雅安市雨城区,平均海拔620 m,属亚热带湿润季风气候区,多年平均气温16.2℃,多年平均降雨量1 743.3 mm,多年平均日照1 035 h,多年平均蒸发量1 011.2 mm。枇杷苗栽种基质采用过2 mm筛的市售建筑用砂,用遮阳网覆盖,每天浇水以保持基质湿润。
少花龙葵嫁接后代种子为2012年收集的备用种子,采用不同砧木对少花龙葵进行嫁接,分别为:不嫁接(UG)、自根同株嫁接(SG)、自根异株嫁接(DG)和野生马铃薯砧木嫁接(PG),本试验采用的是少花龙葵嫁接后代第一代种子(UG1、SG1、DG1和PG1)。2013年6月,将少花龙葵嫁接后代种子直接撒播于四川农业大学雅安校区农场果园(未污染区),待少花龙葵嫁接后代苗高约5 cm(5片真叶展开)时进行移栽。
1.2 试验设计
供试土壤为紫色土,取自四川农业大学雅安校区农场果园,其基本理化性质:pH 7.02,有机质41.38 g/kg,全氮3.05 g/kg,全磷0.31 g/kg,全钾15.22 g/kg,碱解氮165.30 mg/kg,有效磷5.87 mg/kg,速效钾187.03 mg/kg,镉全量0.101 mg/kg,有效态镉含量0.021 mg/kg。土壤理化性质及重金属含量均按照鲍士旦[19]的方法测定。2013年6月,用15 cm × 18 cm (直径×高)塑料盆装入风干土3.0 kg,以溶液形式加入10 mg/kg镉(CdCl2·2.5H2O,分析纯)[18],保持土壤湿润,放置4周,不定期翻土混合,使土壤充分混合均匀。2013年7月,将长势一致的少花龙葵嫁接后代幼苗、两年生大五星枇杷实生幼苗和川早枇杷实生幼苗移栽至盆中,种植处理分别为:UG1单种、SG1单种、DG1单种、PG1单种、川早枇杷单种、UG1混种川早枇杷、SG1混种川早枇杷、DG1混种川早枇杷、PG1混种川早枇杷、大五星枇杷单种、UG1混种大五星枇杷、SG1混种大五星枇杷、DG1混种大五星枇杷、PG1混种大五星枇杷。单种每盆种植植物2株,混种每盆种植每种植物各1株,每个处理重复6次。盆与盆之间的距离为10 cm,完全随机排列。植物生长期间不定期地交换盆与盆的位置以减弱边际效应的影响,并及时浇水,使土壤田间持水量保持在80% 左右。
1.3 测定项目与方法
种植2个月后(2013年9月),对每种植物进行取样测定相关指标。用叶绿素仪(SPAD-502 PLUS,日本产)测定少花龙葵嫁接后代叶片SPAD值(表示叶绿素相对含量),每株测定完全展开的成熟新叶2片(茎尖往下第5、6片叶),测定位置为叶片的中心。选择枇杷幼苗成熟叶片(茎尖往下第2、3片叶)测定叶绿素(叶绿素a、叶绿素b及叶绿素总量)含量,测定方法为丙酮-乙醇混合(,1∶1)浸提法[20]。少花龙葵嫁接后代和枇杷幼苗的光合作用采用LI-6400便携式光合测定仪(LI-COR Inc., USA)测定,选取每株少花龙葵完全展开的成熟新叶2片(茎尖往下第5、6片叶)和枇杷幼苗成熟叶片2片(茎尖往下第4、5片叶),测定位置为叶片的中心,连续测定3天,于每日上午10点开始测定(30 min内测完)。光合作用测定时,人工控制CO2浓度为400 μmol/mol、温度25℃、光照强度为1 200 μmol/(m2·s),测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和叶表面蒸汽压亏缺(Vpdl)。植物根系、茎杆和叶片的可溶性糖含量采用蒽酮比色法测定[20]。
1.4 数据处理
本试验数据采用DPS系统进行方差分析,Duncan新复极差法进行多重比较。
2 结果与分析
2.1 枇杷幼苗光合生理特性
由表1数据可知,混种少花龙葵嫁接后代后,大五星枇杷幼苗和川早枇杷幼苗的叶绿素a含量、叶绿素b含量及叶绿素总量均高于各自单种,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种。混种UG1、混种SG1、混种DG1和混种PG1的大五星枇杷幼苗叶绿素总量较单种分别提高了1.44%(>0.05)、5.75%(<0.05)、10.06%(<0.05)和22.79%(<0.05),而川早枇杷幼苗叶绿素总量较单种分别提高了3.37%(>0.05)、10.71%(<0.05)、14.48%(<0.05)和21.43%(<0.05)。从叶绿素a/b来看,混种少花龙葵嫁接后代后,大五星枇杷幼苗和川早枇杷幼苗叶绿素a/b均高于各自单种,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1 >单种,说明混种后的枇杷幼苗叶绿素a含量所占比重增大。
由表2数据可知,混种少花龙葵嫁接后代提高了大五星枇杷幼苗和川早枇杷幼苗的净光合速率,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种。混种UG1、混种SG1、混种DG1和混种PG1的大五星枇杷幼苗净光合速率较单种分别显著提高了13.76%、19.36%、29.76% 和40.64% (<0.05),而川早枇杷幼苗净光合速率较单种分别显著提高了20.49%、23.91%、34.91% 和36.05% (<0.05)。混种少花龙葵嫁接后代后,大五星枇杷幼苗和川早枇杷幼苗的气孔导度、胞间CO2浓度、蒸腾速率均高于各自单种,但叶表面蒸汽压亏缺均低于各自单种。大五星枇杷幼苗和川早枇杷幼苗的气孔导度、胞间CO2浓度、蒸腾速率的大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种,叶表面蒸汽压亏缺的大小顺序均为:单种>混种UG1>混种SG1>混种DG1>混种PG1。

表1 枇杷幼苗叶绿素含量(mg/g)
注:同列数据后不同小写字母表示同一枇杷品种下不同处理间差异在<0.05水平显著,下同。
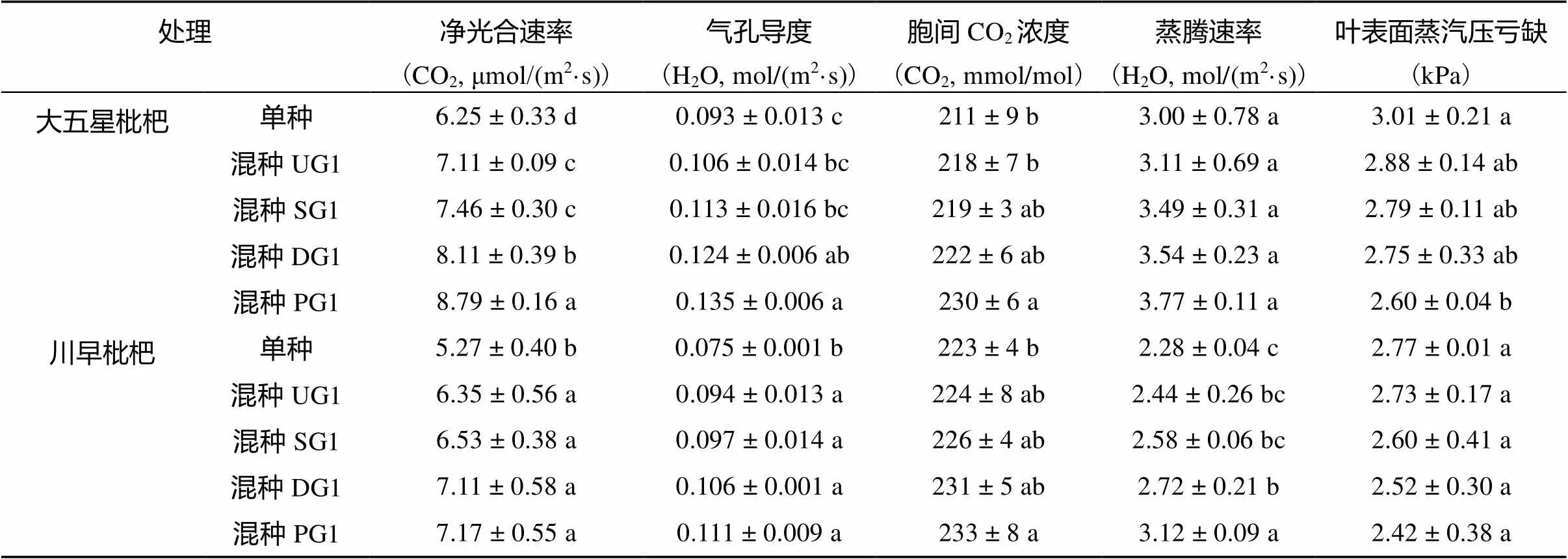
表2 枇杷幼苗光合特性
从表3可得,混种少花龙葵嫁接后代后,大五星枇杷幼苗和川早枇杷幼苗的根系、茎杆和叶片可溶性糖含量均高于各自单种,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种。混种UG1、混种SG1、混种DG1和混种PG1的大五星枇杷幼苗根系可溶性糖含量较单种分别显著提高了12.59%、29.46%、34.35% 和51.45%(<0.05),茎杆可溶性糖含量较单种分别显著提高了22.21%、22.92%、37.12% 和61.82%(<0.05),叶片可溶性糖含量较单种分别显著提高了4.67%、8.44%、10.81% 和21.33%(<0.05)。混种UG1、混种SG1、混种DG1和混种PG1的川早枇杷幼苗根系可溶性糖含量较单种分别显著提高了13.78%、15.91%、20.02% 和23.46%(<0.05),茎杆可溶性糖含量较单种分别显著提高了12.18%、44.84%、51.05% 和54.03%(<0.05),叶片可溶性糖含量较单种分别提高了2.44%(>0.05)、6.65%(< 0.05)、18.24%(<0.05)和18.52%(<0.05)。由此可见,混种少花龙葵嫁接后代后,枇杷幼苗的可溶性糖含量在源(叶片)、流(根系)、库(茎杆)中被重新分配,其中在库中的提高幅度最大,其次为流,源中的最小。
2.2 少花龙葵嫁接后代光合生理特性
从表4可以看出,少花龙葵嫁接后代在单种和混种条件下的净光合速率、气孔导度、胞间CO2浓度及蒸腾速率的大小顺序均为:PG1>DG1>SG1>UG1,但叶表面蒸汽压亏缺的大小顺序均为:UG1>SG1> DG1> PG1。混种枇杷幼苗后,少花龙葵嫁接后代的净光合速率、气孔导度、胞间CO2浓度及蒸腾速率均高于各自单种,且混种大五星枇杷幼苗高于对应的混种川早枇杷幼苗。从总体上看,混种枇杷幼苗后,少花龙葵嫁接后代的叶表面蒸汽压亏缺均低于各自单种,且混种大五星枇杷幼苗高于对应的混种川早枇杷幼苗。
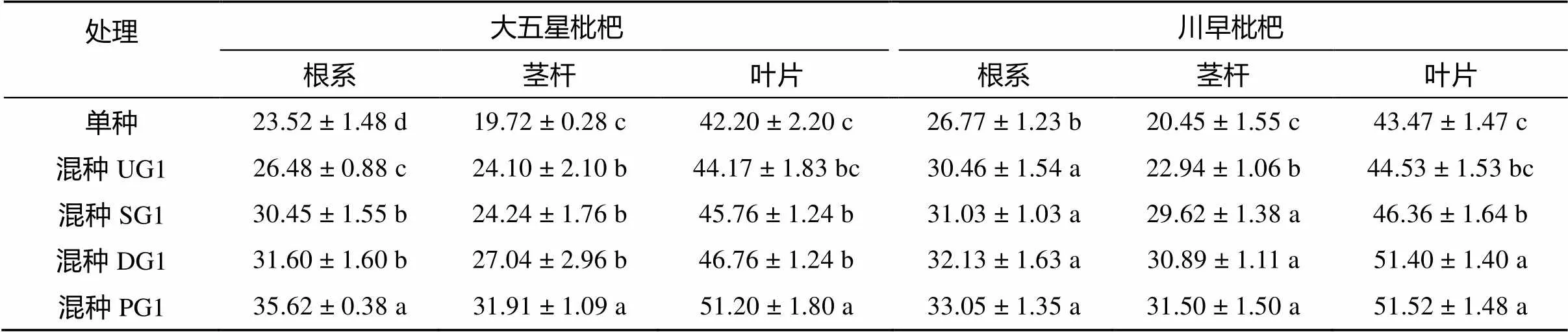
表3 枇杷幼苗可溶性糖含量(mg/g,以干物质量计)
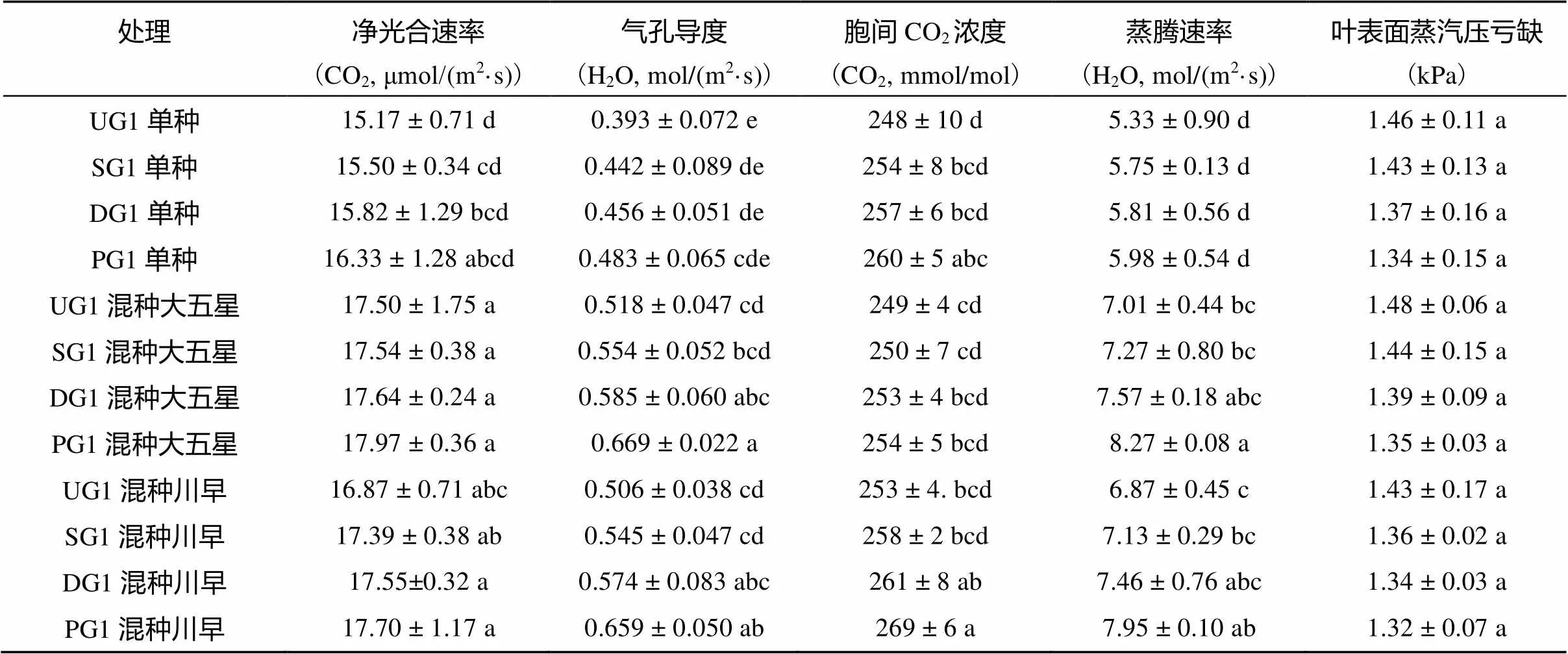
表4 少花龙葵嫁接后代光合特性
从表5可得,少花龙葵嫁接后代在单种和混种条件下的根系、茎杆及叶片可溶性糖含量的大小顺序均为:PG1>DG1>SG1>UG1。与单种相比,混种大五星枇杷幼苗和川早枇杷幼苗的少花龙葵嫁接后代根系、茎杆及叶片的可溶性糖含量均降低,且混种大五星枇杷幼苗均高于对应的混种川早枇杷幼苗,这说明混种调节了少花龙葵嫁接后代植株的可溶性糖代谢。混种大五星枇杷幼苗的UG1、SG1、DG1和PG1根系可溶性糖含量分别较各自单种降低了3.71%(> 0.05)、4.33%(>0.05)、5.31%(<0.05)和4.01%(>0.05),茎杆可溶性糖含量分别较各自单种显著降低了23.18%、17.42%、19.31% 和12.33%(<0.05),叶片可溶性糖含量分别较各自单种显著降低了24.74%、33.94%、39.38% 和38.12%(<0.05)。混种川早枇杷幼苗的UG1、SG1、DG1和PG1根系可溶性糖含量分别较各自单种显著降低了17.36%、19.98%、24.81% 和25.95%(<0.05),茎杆可溶性糖含量分别较各自单种显著降低了26.25%、32.04%、31.34% 和27.17% (<0.05),叶片可溶性糖含量分别较各自单种降低了32.92%(<0.05)、43.61%(>0.05)、48.02% (<0.05) 和51.85%(<0.05)。
从图1可以看出,少花龙葵嫁接后代在单种和混种条件下的SPAD值的大小顺序均为:PG1>DG1> UG1>SG1。混种枇杷幼苗后,少花龙葵嫁接后代的SPAD值均高于各自单种,且混种大五星枇杷幼苗的少花龙葵嫁接后代的SPAD值均小于混种川早枇杷幼苗。这些结果说明混种枇杷幼苗能提高少花龙葵嫁接后代的抗性。
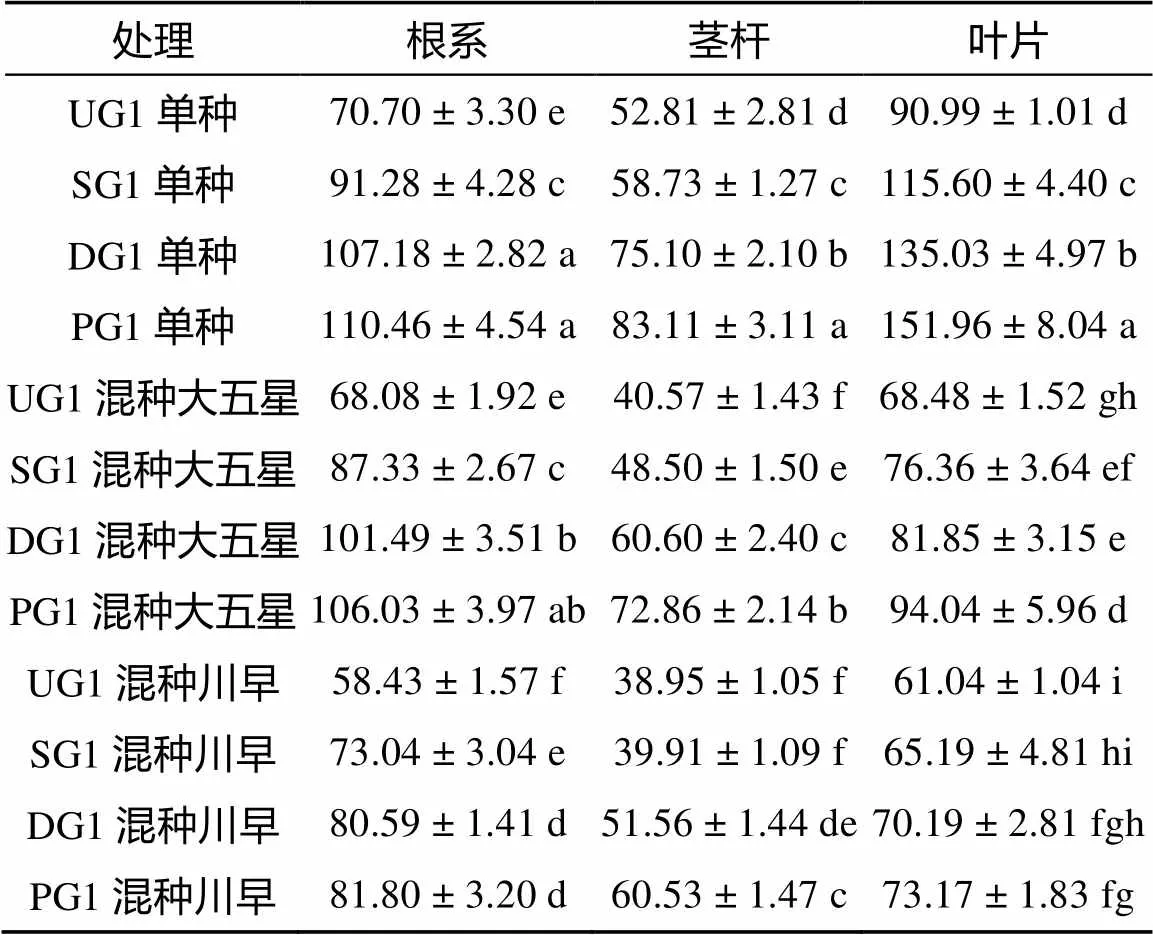
表5 少花龙葵嫁接后代可溶性糖含量(mg/g,以干物质量计)

(图中不同小写字母表示不同处理间差异在P<0.05水平显著)
3 讨论
混种是农业生产的一种常用生产技术,通过长期混种,土壤理化性质能够得到改善,土壤环境中的生物多样性也得到提高,最终作物生长得到促进[21-23],这在玉米间作大豆上表现最为突出[24]。不同植物混种后可以经由“根际效应”、“化感作用”等改善土壤的理化性质,提高土壤环境中的生物多样性,使植物光照利用率増大,光合作用增强,促进植物生长,改善植物生长发育状况。胡举伟等[25]将桑树和苜蓿混种,发现混种苜蓿的叶绿素a和叶绿素b含量都高于单种。杨代宇[26]研究表明,混种鬼针草、万寿菊后树番茄幼苗的生物量、光合色素含量高于单种,但混种少花龙葵、牛膝菊后树番茄幼苗的生物量、光合色素含量都低于单种。本试验研究表明,在镉污染条件下,少花龙葵嫁接后代和枇杷幼苗混种后,枇杷幼苗的光合色素含量均高于其单种,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种。这说明混种在一定程度上促进了枇杷幼苗叶片的水分利用效率,可能与混种后减少了土壤表面水分蒸发有关,因此,少花龙葵嫁接后代适合与枇杷幼苗混种,这种搭配有利于镉污染果园的修复。
混种改变了土壤环境,而根系分泌物是影响植物生长的重要物质[27-28]。在王兴伟等[29]试验中,在重金属污染条件下,用假繁缕和三叶草进行混种后不利于三叶草的地上部分光合及生长,而对假繁缕的生长和光合有利,但假繁缕与黑麦草混种的情况则相反,对假繁缕的光合及生长不利,而对黑麦草有利。黑麦草与其他两种植物混种后,其混种的植物地上部分的生长产生了抑制,三叶草表现出间作劣势,同时给与其混种的植物带来了间作优势,促进了与其混种的植物的地上部分的光合作用。这可能是因为不同的植物在生长过程中分泌的化感物质对土壤养分有效性、酶活性、微生物种群结构等产生了影响,继而调控了与其混种的植物体内的相关生化反应过程,影响光合作用效率,从而表现为对植物地上部生长的促进和抑制两种不同的作用。在光合作用过程中,CO2从空气中向叶片叶绿体光合部位的传播受到众多因素的影响(如细胞内 CO2浓度、气孔导度等),而气孔是植物叶片中最重要的气体运输通道,直接控制了 CO2向叶片的传导以及叶片的蒸腾效率[30]。Wong等[31]研究表明,光合速率与气孔导度之间存在一定的反馈调节机制,在有利于叶肉光合作用时,气孔导度增大;反之则减小。Mersie等[32]研究也表明阿魏酸和香草酸这2种化感物质能导致天鹅绒叶片的气孔导度降低,叶片阻力上升,光合速率显著下降。本试验研究表明,在镉污染条件下,少花龙葵嫁接后代混种枇杷幼苗提高了两者的光合作用,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种。混种少花龙葵嫁接后代后,大五星枇杷幼苗和川早枇杷幼苗的气孔导度、胞间CO2浓度、蒸腾速率均高于各自单种,但叶表面蒸汽压亏缺均低于各自单种,混种枇杷幼苗后,少花龙葵嫁接后代的净光合速率、气孔导度、胞间CO2浓度及蒸腾速率均高于各自的单种,且混种大五星枇杷幼苗高于对应的混种川早枇杷幼苗。在本试验中,少花龙葵嫁接后代和枇杷幼苗均表现出了间作优势,这可能是两种植物的根系分泌物相互刺激,其生理代谢机制重新调整达到一个新的更好的平衡点,进而使其抗性增加,从而促进其光合作用,生长加快。
植物可溶性糖含量的变化是植物碳水化合物代谢的重要指标,它既可反映碳水化合物的合成情况,也与碳水化合物在植物体内的运输和利用情况有关[33],一方面标志着叶源端的同化物供应能力,另一方面也能反映出籽粒对同化物的转化、利用能力[34]。可溶性糖不仅是高等植物的主要光合产物,而且是碳水化合物代谢和暂时贮藏的主要形式,所以在植物代谢中占有重要位置。许多研究结果表明,可溶性糖含量变化与光合作用和产量密切相关[35-36]。Sun等[37]研究小麦的结果也表明,可溶性糖含量变化和光合作用密切相关。本试验研究表明,混种少花龙葵嫁接后代后,大五星枇杷幼苗和川早枇杷幼苗的根系、茎杆和叶片可溶性糖含量均高于各自单种,其大小顺序均为:混种PG1>混种DG1>混种SG1>混种UG1>单种。由此可见,混种少花龙葵嫁接后代后,枇杷幼苗的可溶性糖含量在源(叶片)、流(根系)、库(茎杆)中被重新分配,其中在库中的提高幅度最大,其次为流,源中的最小。少花龙葵嫁接后代在单种和混种条件下的根系、茎杆及叶片可溶性糖含量的大小顺序均为:PG1>DG1> SG1>UG1。与单种相比,混种大五星枇杷幼苗和川早枇杷幼苗的少花龙葵嫁接后代根系、茎杆及叶片的可溶性糖含量均降低,且混种大五星枇杷幼苗均高于对应的混种川早枇杷幼苗,这说明混种调节了少花龙葵嫁接后代植株的可溶性糖代谢。从碳代谢来看,在镉污染条件下,少花龙葵嫁接后代混种枇杷幼苗均降低了少花龙葵嫁接后代根系、茎杆及叶片的可溶性糖含量,提高了枇杷幼苗根系、茎杆及叶片的可溶性糖含量。这说明两种植物的碳代谢均得到了调整,且在其各自的源、流、库中的分配也发生了变化,可能与两种完全不同类别植物(草本和木本)的碳代谢机制不同有关。
4 结论
混种少花龙葵嫁接后代能够提高枇杷幼苗的叶绿素含量和可溶性糖含量,促进其光合作用,同时混种能够提高少花龙葵嫁接后代的SPAD值,但混种枇杷后少花龙葵嫁接后代的可溶性糖含量较各自单种有所降低。
[1] 肖鹏飞, 李法云, 付宝荣, 等. 土壤重金属污染及其植物修复研究[J]. 辽宁大学学报(自然科学版), 2004 , 31(3): 279–283
[2] 陈奕云, 唐名阳, 王淑桃, 等. 基于文献计量的中国农田土壤重金属污染评价[J]. 土壤通报, 2016, 47(1): 219–225
[3] 李江遐, 张军, 黄伏森, 等. 铜矿区土壤重金属污染与耐性植物累积特征[J]. 土壤通报, 2016, 47(3): 719–724
[4] 陈吉美. 镉对植物胁迫效应的研究进展[J]. 安徽农学通报, 2010, 16(3): 46-47
[5] 卢良恕. 中国立体农业概论[M]. 成都: 四川科学技术出版社, 1999: 98–106
[6] 焦念元, 赵春, 宁堂原, 等. 玉米-花生间作对作物产量和光合作用光响应的影响[J]. 应用生态学报, 2008, 19(5): 981–985
[7] 牛斌, 冯莎莎, 张俊平. 小麦玉米玉米间套作的光合变化[J]. 安徽农业科学, 2009, 37(35): 17429–17430
[8] 刘红敏, 宁万光, 徐畅, 等. 不同品种水稻间作栽培对褐飞虱发生及水稻产量的影响[J]. 2015, 43(4): 124–126
[9] 周向军. 重金属超富集植物环境修复技术研究进展[J]. 安徽农业科学, 2010, 38(3): 1408–1410
[10] 王海慧, 郇恒福, 罗瑛, 等. 土壤重金属污染及植物修复技术[J]. 中国农学通报, 2009, 25(11): 210–214
[11] 汤福义, 林立金, 杨代宇, 等. 少花龙葵种间嫁接后代对小白菜生长及镉积累的影响[J]. 土壤通报, 2016, 47(1): 207–212
[12] 杨代宇, 林立金, 张潇, 等. 镉胁迫下不同生态型富集植物混种樱桃幼苗对土壤酶活性的影响[J]. 水土保持通报, 2015, 35(4): 73–78
[13] 吴媛琳, 赵听, 张凯煜, 等. 枇杷不同部位主要有效成分含量及抗氧化活性比较[J]. 西北林学院学报, 2015, 30(1): 196–201
[14] 陈杰忠. 果树栽培学各论(第三版)[M]. 北京: 中国农业出版社, 2003
[15] 余东, 李永裕, 邱栋梁, 等. 镉(Cd)胁迫对枇杷生长和光合速率的影响[J]. 农业环境科学学报, 2007, 26(增刊): 33–38
[16] Zhang X F, Xia H P, Li Z A, et al. Identification of a new potential Cd-hyperaccumulatorby soil seed bank-metal concentration gradient method[J]. Journal of Hazardous Materials, 2011, 189(1/2): 414–419
[17] 李春俭, 马玮, 张福锁. 根际对话及其对植物生长的影响[J]. 植物营养与肥料学报, 2008, 14(1): 178–183
[18] 石军,胡容平,褚旭东,等. 枇杷实生幼苗对镉的积累特性研究[J]. 陕西农业科学,2016,62(6):1–5, 52
[19] 鲍士旦. 土壤农化分析(第三版)[M]. 北京: 中国农业出版社, 2000
[20] 郝再彬, 苍晶, 徐仲. 植物生理学实验[M]. 哈尔滨: 哈尔滨工业大学出版社, 2004
[21] 黎健龙, 涂攀峰, 陈娜, 等. 茶树与大豆间作效应分析[J]. 中国农业科学, 2008, 41(7): 2040–2047
[22] 徐雪风,李朝周,张俊莲. 轮作油葵对马铃薯生长发育及抗性生理指标的影响[J]. 土壤, 2017, 49(1): 83–89
[23] 侯慧,董坤,杨智仙,等. 连作障碍发生机理研究进展[J]. 土壤, 2016, 48(6): 1068–1076
[24] 唐永金, 刘俊利, 郑占, 等. 玉米大豆间混种植对大豆产量和品质的影响[J]. 大豆科学, 2011, 30(6): 954–958
[25] 胡举伟, 朱文旭, 张会慧, 等. 桑树/首猜间作对其生长及土地和光资源利用能力的影响[J]. 草地学报, 2013, 21(3): 494–500
[26] 杨代宇. 混种两种生态型少花龙葵杂交F1代对树番茄幼苗镉积累的影响[D]. 成都: 四川农业大学, 2016
[27] Tatár E, Mihucz V G, Varga A, et al. Determination of organic acids in xylem sap of cucumber: Effect of lead contamination[J]. Microchemical Journal, 1998, 58: 306–314
[28] Shen Z G, Zhao F J, McGrath S P. Uptake and transport of zinc in the hyperaccumulatorand the non-hyperaccumulator[J]. Plant Cell and Environment, 1997, 20: 898–906
[29] 王兴伟, 刘子芳, 赵兵, 等. 铅锌胁迫下混种黑麦草和三叶草对假繁缕生理生态的影响[J]. 陕西农业科学, 2017, 63(01): 30–34.
[30] 应小芳, 刘鹏. 铝胁迫对大豆叶片光合特性的影响[J]. 应用生态学报, 2005, 16(1): 166–170
[31] Wong S C, Cowan I R, Farquhar G R. Stomatal conductance correlates with photosynthetic capacity[J]. Nature, 1994, 282: 424–426
[32] Mersie W, Singh M. Phenolic acids affect photosynthesis and protein synthesis by isolated leaf cells of velvet-leaf[J]. Journal of Chemical Ecology, 1993, 19: 1293–1301
[33] 王芳, 刘鹏, 朱靖文. 镁对大豆游离脯氨酸、可溶性糖和可溶性蛋白质含量的影响[J]. 河南农业科学, 2004(6): 35–38
[34] Saratha K, Hume D J, Godfrey C. Genetic improvement in short season soy beans: I. Dry matter accumulation, partitioning, and leaf area duration [J] .Crop Science, 2001, 41: 391–398
[35] Jin J, Liu X B, Wang G H. Some eco-physiological characteristic R4-R5 stage in relation to soybean yield differing in maturities [J]. Scientia Agricultura Sinica, 2004, 37: 1293–1300
[36] Wilcox J R. Sixty years of improvement in publicly developed elite soybean lines[J]. Crop Science, 2001, 41: 1711–1716
[37] Sun J Y, Wang Q M, Chen J, et al. Characteristics of triose phosphate/ phosphate translocator from wheat and its role in the distribution of assimilates[J]. Acta Botanica Sinica, 2004, 46(3): 294–301
Effects of Intercropping with Post-Grafting Generation ofon Photosynthetic Physiology of Loquat Seedlings Under Cadmium Stress
LI Hongyan1, CHEN Fabo2, HUANG Jiajing3, WEN Keng3, LIAO Ming’an3,LIN Lijin1*, JIANG Wei4, LI Huaxiong5
(1 Institute of Pomology and Olericulture, Sichuan Agricultural University, Chengdu 611130, China; 2 Life Science and Technology Institute, Yangtze Normal University, Chongqing 408100, China; 3 College of Horticulture, Sichuan Agricultural University, Chengdu 611130, China; 4College of Chemistry and Life Science, Chengdu Normal University, Chengdu 611130, China; 5 Institute of Forestry and Pomology, Neijiang Academy of Agricultural Sciences, Neijiang, Sichuan 641000, China)
A pot experiment was conducted to study the effects of intercropping with post-grafting generation ofon photosynthetic physiology of loquat seedlings under cadmium (Cd) stress, in which 4 post-grafting generations ofwere intercropped with two kinds of loquat seedlings (Dawuxing and Chuanzao) in Cd-contaminated soil. The results showed that, compared with the monoculture, intercropping significantly improved chlorophyll a, chlorophyll b and total chlorophyll contents, net photosynthetic rate, transpiration rate, intercellular CO2concentration, stomatal conductance and soluble sugar content of loquat seedling, decreased the vapor pressure deficit on leaf surface. SPAD and net photosynthetic rate, transpiration rate, intercellular CO2concentration, stomatal conductance of the post-grafting generation ofintercropped were higher than those of monoculture, while the soluble sugar content was lower, the vapor pressure deficit on leaf surface of the post-grafting generation ofintercropped with Dawuxing seedling was higher than the that of monoculture, but the early Sichuan loquat was lower. Therefore, the post-grafting generations ofintercropping with loquat could enhance the photosynthesis of these two plant species, thus could promote the growth of them.
Post-grafting generation of; Loquat; Photosynthetic physiology; Intercropping; Cadmium
四川省科学技术厅应用基础计划项目(2016JY0258)资助。
llj800924@163.com)
李红艳(1996—),女,重庆人,硕士研究生,主要从事果树生理生态研究。E-mail:1432350417@qq.com
S616;S66;S344.2
A
10.13758/j.cnki.tr.2019.04.015
