饲料酶中的主效酶与辅效酶
2019-10-16冯定远
■冯定远
(华南农业大学动物科学学院,广东广州510642)
酶制剂应用于动物日粮的复杂性源于饲料原料的复杂性,饲料原料的复杂性主要体现在物理特性和化学特性两方面。物理特性主要是不同成分的空间结构与嵌合方式和程度等;化学特性主要是同一类成分的组成单位的差异与主链长短及支链分布形式等。这种复杂性既表现在营养成分之间(如蛋白质、淀粉、脂肪间的嵌合),也表现在同一类营养成分内部(如蛋白质中氨基酸的排列组合、淀粉中抗性淀粉的差异、脂肪中脂肪酸1、3 位点与2 位点的差别等),更表现在营养成分与抗营养成分的嵌合与影响,这决定了酶制剂在日粮中应用不简单,单酶或者普通的复合酶往往受制于这些复杂性而不能很好地发挥作用,我们提出了组合酶、配合酶、组合型复合酶和组合型配合酶,就是针对这类饲料日粮常常出现的问题。但是,这里还有一个问题没有解决,就是组合酶或者配合酶中,各种单酶之间的关系问题。另外,不同来源酶的作用底物也存在差异性,如催化作用的位点和结合键合方式的不同的影响等。实际上。复杂的成分与多样的酶相互作用存在一种主次关系、主从关系、主要与辅助关系。自然环境产生的酶,大部分是以一个酶系、整个酶族群存在,这种酶系或者酶群一般是有关联的、有序的,之间的重要程度是有区别的。过去提出组合酶和配合酶并没有讨论到这些关系,更多是看作等同与平衡关系。其实,普通的复合酶作用同样也存在哪个为主,哪个为辅的问题。这也是为什么目前的复合酶配制五花八门,酶谱多样的一个原因。所以,有必要探讨各种单酶之间的关系,明确是哪个酶发挥更大作用,在复合酶、组合酶或者配合酶的研发中兼顾高效性与经济性。
1 饲料主效酶与辅效酶的依据
1.1 物料物理结构的序列性
纤维素是由葡萄糖通过β-1, 4糖苷键链接而成的线状结构分子,具有简单的初级和复杂的三级结构。纤维素链的聚合度(葡萄糖残基数)从500到14 000不等(Marx-Figini等,1966)。植物细胞壁中,纤维素链的排列有序程度不同,在一些区域,纤维素链高度有序排列,在氢键作用下形成晶体;而其他区域则松散排列,纤维素分子形成非晶体状。天然晶体纤维素的结构为I 型,通过碱处理可以转化成Ⅱ型结构(Beguin 等,1994),这两种类型的纤维素的区别在于链内氢键间存在差异。此外,天然结晶纤维素可能由两种结构稍有差异的I型纤维素构成,即Iα、Iβ,这两种亚型的区别也在于其分子内氢键间有差异(Atalla等,1984)。
纤维素中的晶体化部分僵硬,不易被内切作用的纤维素酶降解,而非晶态的纤维素则易被稀酸、内切葡聚糖酶、外切葡聚糖酶降解(Sinitsyn 等,1990)。因此,完全降解纤维素需要采用浓酸或复合酶系,攻击其结晶区和非结晶区。
使用高纯度的内切葡聚糖酶和来自嗜松青霉(P.pinophium)的CBHS的试验,只有二种内切葡聚糖酶(EG Ⅲ和EGⅤ)对纤维素具有很强的吸附性,与CBHSⅠ和CBHSⅡ有协同作用(见图1,Wood 等,1989)。作者认为在CBHSⅠ和Ⅱ与内切葡聚糖酶之间的协同作用,是因为这两类酶间的不同立体空间结构造成的。所以物理空间的不同,会造成酶水解的差异。有特异立体结构的内切葡聚糖酶催化纤维素主链,只可产生一到两种非还原性残基末端,后者可被有特异立体结构的CBH水解。通过CBH连续水解纤维二糖的同时,将暴露另一个不同特异立体结构的链端,会受到其他具有特异立体结构的CBH催化分解。
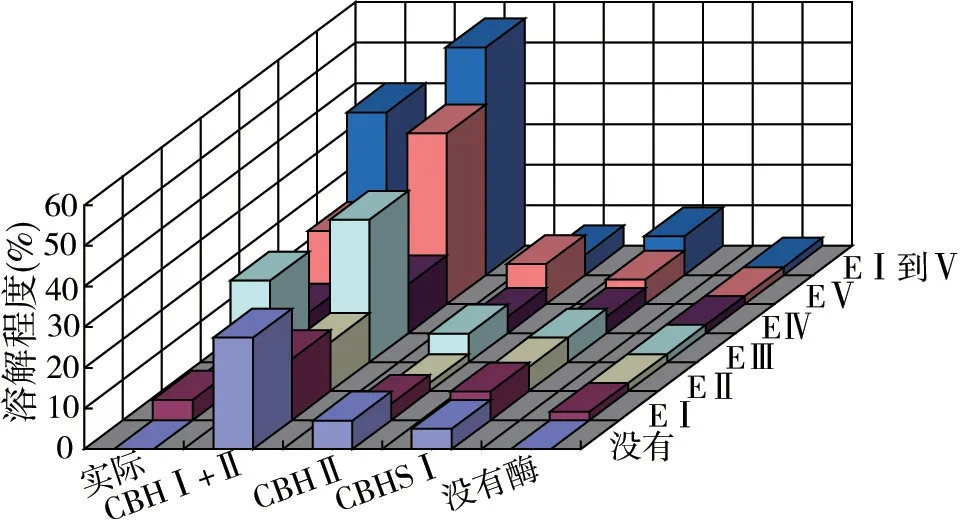
图1 来自嗜松青霉(P. pinophlum)的CBHS(Ⅰ和Ⅱ型)与内切葡聚糖酶(EⅠ至EⅤ)之间在水解棉花纤维上的协同作用
在CBH和CBH(外切葡聚糖酶和外切葡聚糖酶)之间的协同作用,主要是因为其相互为对方提供新的水解结合部位的能力,以及从还原端到非还原端的催化能力(Barr 等,1996)。大量的研究表明:在聚合的嗜热纤维单胞菌(C. thermocellum)酶系中,酶复合体的组装对于在结晶纤维素的溶解中亚基间的最大催化协同效应是十分关键的(Bhat 等,2001)。看起来简单的棉花纤维素就如此复杂,饲料物料就更不用讲了。从仿生角度,人工配制的酶制剂添加剂必须选择不同酶进行配合,才能达到高效水解的目的。
1.2 底物化学成分的差异性
木聚糖(Xylan)是植物细胞壁的重要组成成分,连接细胞壁中的木质素和纤维素,是最主要的半纤维素,广泛存在于硬木(15%~30%)、软木(7%~10%)和草本类植物中(低于30%)。两种主要的阿拉伯木聚糖是:①来源于种子胚乳的高度分支、且没有糖醛酸取代基的阿拉伯木聚糖;②存于木质化组织中的、有较少分支、有糖醛酸和/或4-氧-甲醚以及半乳糖取代基的阿拉伯木聚糖(Bhat等,2001)。木聚糖的主链由木糖经β-1,4-糖苷键连接而成,而其支链结构由于来源的不同而有所差异。
木聚糖具体分类有:①葡萄糖木聚糖(glucuronoxylans);②阿拉伯糖木聚糖(arabinoxylans);③葡萄糖阿拉伯糖木聚糖(glucuronoarabinoxylans),都是以五碳糖的木糖以β-1, 4 键连接形成聚糖作为主链骨架,侧链上是葡萄糖、阿拉伯糖或者同时有葡萄糖和阿拉伯糖(Rogowski等,2015)。第一种的代表是桦木来源的木聚糖,第二种的代表是小麦来源木聚糖,第三种的代表是玉米来源木聚糖。玉米的木聚糖主要是葡萄糖、阿拉伯糖木聚糖,同时含有葡萄糖和阿拉伯糖,而且支链特别多,更复杂。所以,一般的木聚糖酶未必能够发挥作用。谷物类饲料中(小麦、玉米、燕麦)木聚糖的存在形式主要为阿拉伯木聚糖(AX),即主要的支链结构为阿拉伯糖取代基;而来源于硬木(桦木、榉木等)的木聚糖的支链结构则主要为乙酰基、葡萄糖醛酸等。阿拉伯糖主要在主链的C2 或C3位置进行双取代或单取代,而乙酰基主要在C3 位置进行单取代(Kormelink 等,1993)。除这两种侧链取代基外,木聚糖的支链结构还包含多种支链,如阿魏酸(ferulic acid)、葡萄糖醛酸(glucuronic acid)、香豆素(p-coumaric)等(Gruppen 等,1992)。木聚糖的侧链取代基,包括阿拉伯糖和阿魏酸,能够对木聚糖酶发挥空间阻力作用(Akin等,2008)。
1.3 天然存在的酶系与酶之间的协同作用
饲料酶制剂针对的底物包括三类:营养性底物、抗营养底物和功能成分前体物,在自然状况下,都不同程度存在酶系协同作用的现象,例如,简单的脂肪水解,除了广谱性的脂肪酶催化外,更多的是催化甘油1、3 位点与催化2 位点脂肪酸的脂肪酶协同完成。而饲料最复杂的成分是纤维部分,包括纤维素与半纤维素(有六碳糖和五碳糖,其中重要的是木聚糖),为了讨论的方便,我们聚焦于催化纤维素和木聚糖的纤维素酶和木聚糖酶,两种重要的非淀粉多糖酶。我们在这里引用Bhat等(2001)的分析讨论。
纤维素酶和木聚糖酶由细菌和真菌产生,如需氧性微生物(aerobes)、厌氧性微生物(anaerobes)、嗜温微生物(mesophiles)、嗜热微生物(thermophiles)和极温微生物(extremophiles)。好氧性真菌和细菌通常产生细胞外的纤维素酶和半纤维素酶。
厌氧性细菌[热纤维梭菌(Clostridium thermocellum)、嗜纤维梭菌(C. cellulovorans)、白色瘤胃球菌(Ruminococcus albus)、生黄瘤胃球菌(R. flavefaciens)、产琥珀酸丝状杆菌(Fibrobacters succinogenes)、解纤维素醋弧菌(Acetivibrio celluolyticus)]和厌氧性真菌[(Neocallinmastix frontalis)、(N. patriciarum)、(Piromyces equi)]均以多酶复合体的形式产生纤维素酶(Groleau 等,1981;Lamed 等,1987;Wood,1992a;Gilbert等,1993;Beguin等,1996;Bhat等,1997)。
内切葡聚糖酶能特异地作用于非晶态的、膨胀的、取代的纤维素和纤维木聚寡糖内的β-1,4 糖苷键。这些内切葡聚糖酶通常对结晶纤维素和纤维二糖不敏感,可攻击大麦葡聚糖的β-1,3 键和β-1,4 键(Petre 等,1986)。纤维二糖水解酶对结晶纤维素和纤维寡糖的降解能力有限(Wood等,1988),但对非晶态的、膨胀的纤维素有较高活性。自然界中有两种类型纤维二糖水解酶:优先从还原端降解纤维素链和特异性地从非还原端释放纤维二糖(Barr 等,1996;Teeri,1997)。
内切木聚糖酶又可分为特异性和非特异性木聚糖酶(Coughlan,1992;Coughlan 等,1993)。特异性内切木聚糖酶仅对木聚糖的β-1,4键有活性,而非特异性内切木聚糖酶可以水解以β-1,4键连接的木聚糖、混合木聚糖的β-1,4键及其他β-1,4连接的多糖。
大多数内切木聚糖能特异性地作用于木聚糖的非取代木糖苷键,并释放取代的和非取代的木寡糖。相反,其他内切木聚糖酶特异性作用于在主链上的接近取代基团的木糖苷键。例如,来源于黑曲霉(Aspergillus niger)的两种酶(PI 8.0 和9.6)对去掉阿拉伯糖取代基的木寡糖和木聚糖表现很小活性或没有活性(Frederick等,1985)。
Giligan等(1954)在水解纤维素的过程中,首次证实了不同纤维素酶间的协同增效作用。Bhat等(2001)总结了真菌纤维素酶中存在五种协同作用:①内切葡聚糖酶和一种称为C1的非水解蛋白间的协同作用(Reese 等,1950);②β-葡木糖酶和内切葡聚糖酶或者CBH 之间的协同作用(Eriksson 等,1985);③二个免疫学上相关的或截然不同的CBH 之间的协同作用(Wood等,1986);④源自相同或不同微生物内切葡聚糖酶和CBH之间的协同作用(Wood等,1989);⑤二种内切葡聚糖酶之间的协同作用(Klyosov,1990)。另外,细菌纤维素酶和真菌纤维素酶之间的协同作用,与来自嗜热纤维素单胞菌(C. thermocellum)的多酶复合体的亚基之间的协同作用一样(Bhat 等,1994;Wood 等,1994)。协同模式有:①内切葡聚糖酶和外切葡聚糖酶间(CBH)的协同作用;②外切葡聚糖酶和外切葡聚糖酶之间的协同作用;③内切葡聚糖酶和内切葡聚糖酶之间的协同作用。
曲霉菌(Aspergillus)和隐酵母(Cryptococcus)也能产生木聚糖酶和木聚糖去侧链酶(Coughlan 等,1993a,b;Viikari 等,1993)。厌氧微生物能产生降解木聚糖、甘露聚糖的复合多酶系统,这是厌氧微生物的特征(Beguin 等,1996;Hazlewood 等,1998b)。与纤维素类似,有效而彻底的分解木聚糖需要有不同特性的主链裂解酶和支链裂解酶的协同作用(Coughlan等,1993;Coughlan 等,1993)。分解木聚糖的酶协同作用模式也有三种类型:①同型协同;②异型协同;③抗协同(Coughlan等,1993)。同型与异型协同可能有一种或二种产物。同型协同可以是在二种或多种的侧链裂解酶之间的协同,也可能是在二种或多种的主链裂解酶之间的协同(Coughlan等,1993)。
一个同型协同的例子是从米曲霉(A. oryzae)中提取阿魏酸酯酶和从胶囊青霉(P. capsulatum)中提取的α-L-阿拉伯呋喃糖苷酶之间的协同作用,通过后
者使得前者水解阿魏酸化的阿拉伯木聚糖,释放阿拉伯糖变得容易(Coughlan等,1993)。异型协同作用是一种在支链和主链裂解酶间的相互协同作用(Coughlan等,1993)。如果主链裂解酶的作用使得支链裂解酶释放的取代基更多,那么这种异型协同作用有唯一产物,反之亦然。如果通过复合酶的作用,取代基的释放量和主链的水解程度,超过了单一酶制剂的添加作用,则此异型协同作用是个双产物过程。
在阿魏酸酯酶和内切葡聚糖酶之间(Faulds 等,1991;Tenkanen 等,1991)、α-L-阿拉伯呋喃糖苷酶与内切葡聚糖酶之间(Tuohy等,1992),乙酰木聚糖酯酶和内切葡聚糖酶之间(Biely 等,1986;Lee 等,1987),以及葡萄糖苷酸酶和内切葡聚糖酶之间均有异型协同作用。
当一种类型酶的作用阻碍另一种类型酶的作用时,就会出现抗协同作用(Coughlan 等,1993)。这在阿拉伯木聚糖水解酶(Fredeick 等,1985)或者与葡聚木聚糖葡聚糖水解酶(Nishitani 和Nevins,1991)的作用中已有表现,一种酶仅在附近有特殊取代基时,才能裂解主链。葡聚木聚糖葡萄糖苷酸水解酶取代基的存在是其对主链裂解所必需的,通过相关的侧链裂解酶对取代基裂解后,葡聚木聚糖葡萄糖苷酸水解酶将不能裂解主链。
2 饲料主效酶及辅效酶的概念和意义
Bhat 等(2001)在专门讨论“纤维素酶和木聚糖酶酶学及其他特征”的综述时就总结指出:仍有一些问题值得探讨“纤维素酶和半纤维素酶形成多酶复合体和解聚合的相对效率;在水解复杂木质纤维素底物过程中单一酶的作用”。从这两位专家提到的“多酶复合体”中各种单酶存在相对效率,我们很自然就推导到相互之间有主次关系、从属关系。最近Cozannet 等(2017)也研究了“多碳水化合物酶复合体,multi-carbohydrase complex,MCC”中木聚糖酶与阿拉伯呋喃糖苷酶的协同作用。
为了彻底水解复杂的物料,天然的微生物能够产生一个相互配合、协同作用的酶系,一个相关联的酶(族)群,或者称为“多酶复合体”。过去我们讨论的“组合酶”和“配合酶”就属于多酶复合体,其中组合酶是针对同一类底物,例如多种纤维素酶的组合,配合酶是针对不同类底物,如纤维素酶与木聚糖酶的配合(冯定远等,2011)。在这种多酶复合体中,各种酶的关系,特别是平衡关系、主次关系、从属关系如何?相对效率如何?是否可以量化等等?并没有引起注意。
探讨这个问题,我们可以提出一个概念,就是主效酶与辅效酶,或者是基础酶与附属酶的关系。
所谓“主效酶”就是:在多酶复合体中,针对目标底物起基本作用,发挥主导功能,主攻催化化学构成中主链位点或者物理空间主体成分的单酶。相对应,“辅效酶”是指在多酶复合体中,发挥辅助性,次要性甚至是依附性作用,在某些侧链或者支链(一般比侧链长)起作用的单酶。
“主效酶”的另一个理解是单独使用时,其达到目标的效率比“辅效酶”更高。但是,主效酶与辅效酶配合作用,应起到协同的效果,比各自单独使用的效率更高。
实际上,“主效酶”与“辅效酶”是相对概念,在不同的催化水解目的中,是可以互换的。例如,在纤维素、半纤维素及其产物中,有时候是抗营养因子,有时候是益生元的原料,简单理解,木聚糖酶对消除木聚糖的黏性是主效酶,α-L-阿拉伯呋喃糖苷酶和阿魏酸酯酶等侧链糖苷酶可能是辅效酶,相反,如果是产生益生元调节肠道菌群,情况可能不一样。
“主效酶”和“辅效酶”可以体现于组合酶、配合酶这类关联度紧密的酶之间关系中,广义上理解,普通的复合酶也可能存在主效与辅效的关系,只是这种主次关系并没有从属关系。例如在含有蛋白酶和木聚糖酶等的复合酶中,针对小麦型日粮,木聚糖酶是主效酶,而蛋白酶可能是辅效酶,针对杂粕型日粮,蛋白酶往往扮演主效酶的角色,木聚糖酶则是辅效酶。
一般地,主效酶是一个单酶,辅效酶可以有多个不同的单酶。当然,单一纯合的底物不需要辅合酶。
与主效酶和辅效酶相关联的一个容易辩识的概念是底物成分的主链和侧链及相应的主链酶和侧链酶,严格上讲,并没有主链酶和侧链酶的讲法,这是一个通俗的称谓。另外,是侧链还是支链也没有严格的区别,一般是支链更长些,但比主链短,而侧链的基本构成单位更少。
主效酶用一个简单而直接的说法,就是在设计配制特定的饲料酶制剂时,第一个想到的酶,或者重点考虑的酶,就是主效酶。主效酶和辅效酶并不仅仅存在于组合酶和配合酶,这些典型的“多酶复合体”这类关联紧密的单酶之间。如果把日粮配方作为一个整体看,以动物的生产性能作为判断,复合酶中不同的单酶也存在主效酶与辅效酶之别。
在针对一个复杂原料或者有关联的一组有机物成分进行降解的天然存在的酶,往往是一个多种酶的集群,单一的酶作用不大。酶系或者“多酶复合体”应是大自然的普遍情况,正如指出,需要探讨其中单酶的相对效率问题,我们在此基础上,提出“饲料主效酶与辅效酶”的理念。有一定的理论意义及实践价值。
降解复杂木聚糖(AX)的主效酶是木聚糖酶,木聚糖酶也是最广泛应用的饲料添加剂中的酶制剂之一(冯定远,2013)。Paloheimo 等(2010)深入讨论了以饲料添加剂形式的木聚糖酶。他认为在饲料酶制剂市场中,植酸约占一半,其余基本是非淀粉多糖酶(NSP酶),而NSP 酶则最多的是木聚糖酶。木聚糖酶种类很多,黑曲酶(A.niger)和绿色木霉(T. viride)就可分别产生15种和13种木聚糖酶(Biely,1985)。酶蛋白的氨基酸序列和空间结构的不同,决定了它们在水解木聚糖时的酶切位点存在差异性,因而根据水解糖苷键的不同,木聚糖酶可分为不同的糖苷水解酶家族(glycoside hydrolase family,GH),如GH5、GH7、GH8等(Coughlan 等,1993)。用作饲料添加剂的一般是糖苷水解酶家族的GH10 和GH11 的木聚糖酶(Paloheimo 等,2010)。常见的饲用Xyn属于糖苷酶家族10(作用于主链上的≥2个木糖连接体)和糖苷酶家族11(作用于主链上的≥3 个木糖连接体),分解主链木聚糖上的木糖苷键,属于内切酶。植物中的木聚糖结构,一般每10 个木糖单位中有8~9个带有侧链结构,阻碍Xyn与主链结构的接触,使Xyn不能有效发挥作用,只有充分降解木聚糖的侧链,Xyn的作用功效才会增加。
木聚糖(AX)的完全降解需要多种酶的协同作用才能完成(Collins等,2005),主要包括内切型木聚糖酶(endo-1,4-β-D-xylannase, EC 3.3.1.8)、木糖苷酶(β-D-xylosidases, EC 3.2.1.37)以及木聚糖支链分解酶(accessory enzymes),包括α-L-阿拉伯呋喃糖苷酶(α-L-arabinofuranosidase, EC.3.2.1.55)、乙酰木聚糖酯酶(acetyxylan esterase, EC 3.1.1.72)、阿魏酸酯酶(ferulic acid esterase, EC 3.1.1-)等。而作为复杂木聚糖降解的辅效酶阿拉伯呋喃糖苷酶(Abf)和阿魏酸酯酶(FAE)则被了解不多。阿拉伯呋喃糖苷酶和阿魏酸酯酶分别可以特异性地降解AX支链的阿拉伯糖和阿魏酸酯键(Poutanen 等,1988),从而利于木聚糖酶对AX的降解作用(Akin等,2008)。AbfB能够催化多聚糖、低聚糖和多糖非还原端α-连接的阿拉伯残基的水解(陈芳芳,2018),生成一个α-L-阿拉伯糖分子。当AbfB 和Xyn共同作用于AX时,首先侧链的阿拉伯糖快速被降解而释放,然后Xyn才作用于主链木聚糖释放木糖;阿拉伯糖的移除给Xyn 提供了作用位点,同时也增加了底物的溶解性。因此,AX的降解,首先要移除侧链,然后再降解主链。研究表明,在半纤维素酶系中额外添加AbfB,可以明显增加还原糖的生成量(Huang,2017)。而FAE能水解主链中的阿魏酸酯键,释放出阿魏酸或阿魏酸二聚体(范韵敏,2012)。当FAE 和Xyn 共同作用于AX 时,首先侧链的阿魏酸酯键快速被FAE 破坏,为Xyn 提供了合适的作用位点,然后Xyn才作用于主链木聚糖释放木糖。
添加阿拉伯呋喃糖苷酶和阿魏酸酯酶都可以显著提高木聚糖酶降解AX的协同因子(degree of synergistic value),促进AX的降解,生成更多的还原糖(Lei等,2016)。阿拉伯呋喃糖苷酶(McCleary等,2015)和阿魏酸酯酶(Wong 等,2013)同木聚糖酶的协同作用分别在谷物副产物上也得到了证实。此外,先采用支链降解酶去除AX的支链结构,再用木聚糖酶降解AX的 效 果 会 更 好(Raweesri 等,2008;Sorensen 等,2007)。阿拉伯呋喃糖苷酶和阿魏酸酯酶的同时添加,能够较单独添加两种支链酶,进一步提高木聚糖的降解作用(Lei等,2016)。
饲料主效酶与辅效酶提出的意义有多个方面:一是深化对单酶催化作用的研究,也利于深入了解酶作用的机理,酶发挥作用位点,障碍酶催化的影响因素等;二是探讨酶之间的协同性、抗协同性及其生化基础;三是确定主效酶是酶制剂添加剂的基础,主效酶往往可以以单酶形式作为饲料添加剂,例如:植酸酶、木聚糖酶、蛋白酶常常是以单酶形式在日粮中添加。四是在日粮结构改变并更复杂的情况下,饲料酶制剂酶谱设计需要相适应,在确定主效酶的情况下,再增加不同的辅效酶;五是确定了主效酶与辅效酶以及有多少个辅效酶后,在配制饲料酶酶谱,或者选择饲料酶产品时,可以根据成本因素,灵活多样,也就是所谓的产品的“性价比”更有依据;六是复杂的饲料原料与多酶复合体的关系研究,在确定了主效酶与更多个辅效酶的关系后,更有利于开发新的饲料资源。
3 饲料酶制剂中主效酶与辅效酶的含义与应用实践(见图2)
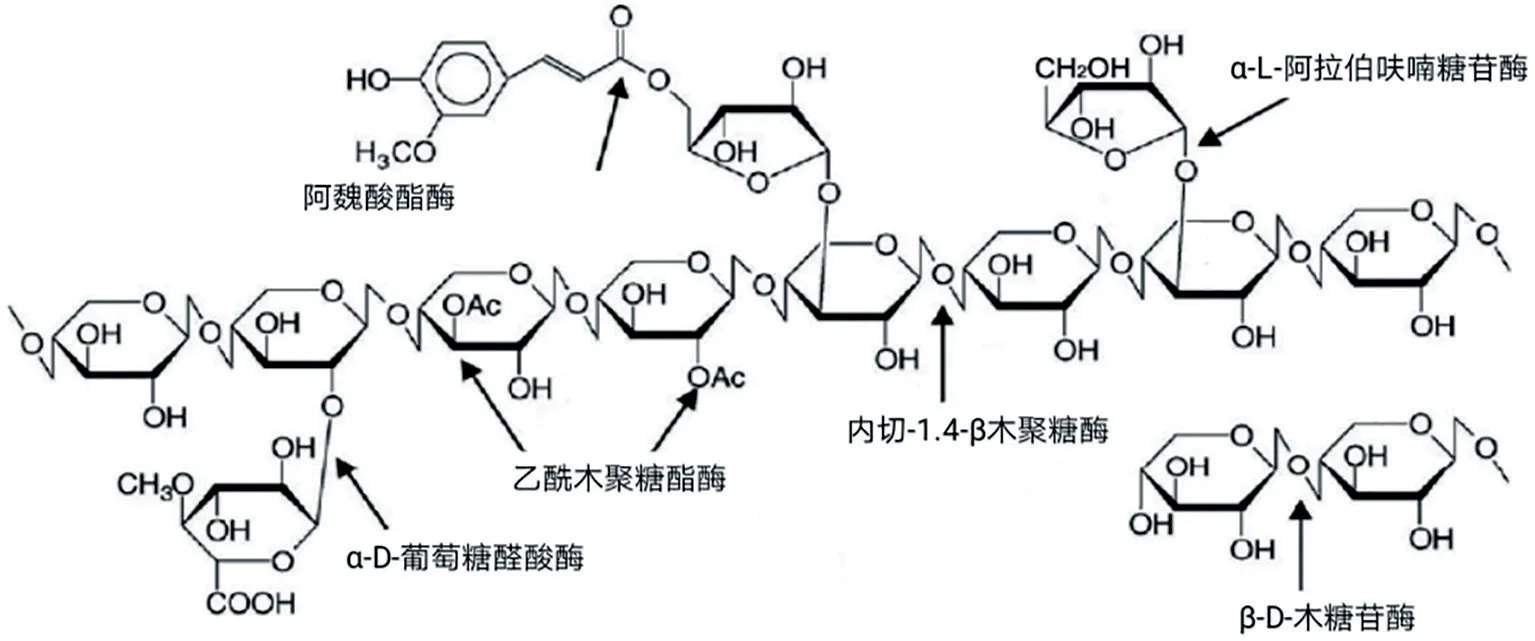
图2 复杂木聚糖的主链与侧链及相应酶作用位点
上面已讨论了自然条件下,纤维素酶和木聚糖酶的复杂性,一般是以酶系或者“多酶复合体”形式存在而彻底水解纤维类物料。如果在配制的饲料酶中,模拟“多酶复合体”而选择主效酶与辅效酶最好的模型就是带有侧链或者支链的木聚糖。在谷麦类饲料原料中,木聚糖的主要存在形式为阿拉伯木聚糖(AX)。AX 以β-1,4-糖苷键连接而成的D-吡喃木糖为主链骨架,在α-1,2 和α-1,3 位由α-L-阿拉伯呋喃糖以侧链的形式进行单取代或双取代,在阿拉伯呋喃糖的O-5位通常会有阿魏酸形成酯键。完全降解AX需要木聚糖酶(Xyn)、阿拉伯呋喃糖苷酶(AbfB)、阿魏酸酯酶(FAE)等若干种酶的协同作用(Collins,2005),形成一个复杂的降解系统:Xyn 作用于主链上,是当中首要的降解酶,随机将长链的木聚糖水解为短链的低聚木糖AX(雷钊,2017);AbfB 则快速移除侧链上的阿拉伯糖,更好地为Xyn 提供作用位点,同时提升了底物的溶解度(解西柱,2018);FAE破坏阿拉伯呋喃糖O-5位形成阿魏酸酯键,简化AX结构,暴露出更多的作用位点(Rahmani,2018)。
降解木聚糖,首先考虑的是木聚糖酶,我们可以把木聚糖酶理解或者设定为主效酶。但是,复杂的侧链影响了一般的木聚糖酶的效率,这时候,就需要考虑辅效酶,这里是侧链酶的阿拉伯呋喃糖苷酶(AbfB)和阿魏酸酯酶(FAE)。关于AbfB 和FAE 并不如木聚糖酶(Xyn)为人们所熟知。侧链上含有阿拉伯残基的木聚糖很难直接被Xyn所降解,主要由于其复杂的支链结构增强了它的抗降解性。Nghiem(2011)的研究表明,针对玉米,单一使用Xyn,只能降解不到2%的木糖,当协同添加FAE和Xyn,可以降解44%的木糖,表明FAE 是一个很重要的支链降解酶。国外已经把FAE 应用在饲料行业,研究表明,在青贮饲料中添加FAE,发现对消化率有明显的提高。王林林(2015)将被FAE酶解过的麦糟复配至肉鸡日粮中,发现肉鸡日增重有提升的同时,料肉比降低了7.63%。
小麦和玉米的NSP主要是阿拉伯木聚糖,分别达干物质的7.3%和4.7%(Bach Knudsen,2014)。简单的木聚糖能够被木聚糖酶水解,然而,复杂的阿拉伯糖替代方式降低了木聚糖酶的效率,特别是玉米及其副产品的情况(Bach Knudsen,2014)。阿拉伯呋喃糖苷酶(Abf)可以在木糖主链骨架上切除阿拉伯糖,为木聚糖酶作用提供方便(De la Mare 等,2013),最终提高了木聚糖酶的效率。尽管水解木聚糖并没有直接提供更多的能量,但由于消除了木聚糖的抗营养性而提高了日粮的消化效率(Wiseman,2000;McCracken等,2002)。
为了研究主链酶(木聚糖酶)和侧链酶(阿拉伯呋喃糖苷酶及阿魏酸酯酶)的配合作用,探讨其协同效果,作者学生郑达文(2019)进行了相关试验,实质就是主效酶与辅效酶协同理念的实践。为了进一步探讨主效酶与辅效酶的含义与关系,我们这里以广泛存在于谷物中复杂木聚糖的解决方案中使用的多酶复合体为例。
我们的试验(郑达,2019)选取两种常见高纤维原料小麦麸皮和燕麦麸皮,其水可溶性膳食纤维SDF和水不可溶性膳食纤维IDF,分析复杂木聚糖降解的辅效酶,即两种新型支链降解酶:阿拉伯呋喃糖苷酶(AbfB)和阿魏酸酯酶(FAE)的酶学特性,并作用在不同溶解度的膳食纤维DF上,以体外仿生消化酶解试验,找出木聚糖酶(Xyn)与AbfB、FAE的最佳协同配比,并通过动物试验验证在饲料中添加新型复合酶对断奶仔猪生长性能及肠道环境的改善效果。动物试验分为三个处理组,空白对照组饲喂不添加外源酶的基础日粮,酶A和酶B组在饲喂基础日粮的基础上按400 g/t的剂量添加酶制剂A(含木聚糖酶、α-淀粉酶、蛋白酶、甘露聚糖酶)和酶制剂B(木聚糖酶、α-淀粉酶、蛋白酶、甘露聚糖酶、阿拉伯呋喃糖苷酶和阿魏酸酯酶)。
在添加主效酶Xyn 的基础上额外分别加入辅效酶AbfB或FAE,HPLC法和DNS法测定还原糖的结果如图3 所示,还原糖总释放量能提高11%或10.5%,Xyn 与AbfB 或FAE 最佳添加比例为3 000:1 或500:1,AbfB 或FAE 均能提高Xyn 的切割效率,对于还原糖的总体释放量有显著的提高作用。通过提取得到纯度达95%的不同溶解度的DF,并按最佳添加比在DF中加入Xyn和AbfB或FAE,还原糖生成量显著提高(P<0.05)、干物质消化率和能量消化率均明显提升的趋势(0.05<P<0.1)(郑达文,2019)。
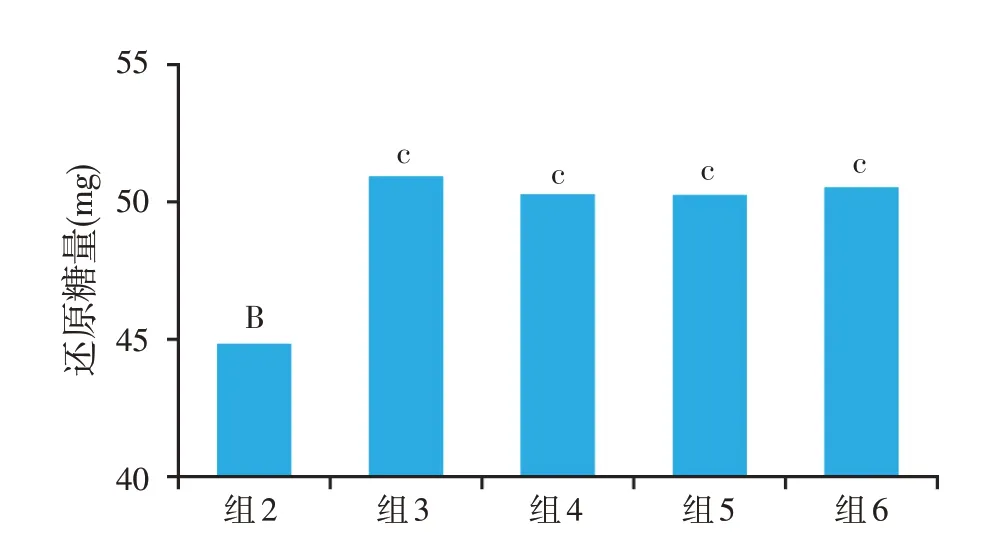
图3 Xyn与AbfB处理淀粉小麦麸产生还原糖的协同作用
SDS体外仿生消化仪评估显示,对比常规外源酶A组和空白对照组,额外添加AbfB或FAE的酶B组的各项表观消化指标均有提升的趋势(0.05<P<0.1),其中:DM%、CP%、Ash%、GE%分别提高0.49%和3.44%、0.57%和3.62%、0.54%和3.71%、0.67%和3.79%。
由表1 可知,对比常规外源酶A 组和空白对照组,额外添加AbfB 和FA 的酶B 组的各项生长性能指标均有较明显的提升趋势(0.05<P<0.1),仔猪ADG、ADFI分别提高3.93%和11.39%、10.77 g/d和18.31 g/d,FCR 和腹泻率则分别下降1.73%和7.52%、0.38%和0.7%,其中腹泻率表现出显著下降(P<0.05)。额外添加阿拉伯呋喃糖苷酶和阿魏酸酯酶的B 组比只添加木聚糖酶的A 组,平均日增重从377.62 g/d 提高到391.43 g/d,有改善效果,但未达到显著水平,而减少腹泻率方面则有显著效果(郑达文,2019)。
另外,加酶组盲肠内容物中乙酸、丙酸、丁酸和总VFA 含量均比空白对照组表现出升高的趋势(0.05<P<0.1)或显著提升(P<0.05);额外添加AbfB、FAE 的酶B 组比常规外源酶A 组,丙酸呈现升高的趋势(0.05<P<0.1),乙酸、丁酸和总VFA 含量极显著提升(P<0.01)。在结肠内容物中,加酶组的乙酸、丙酸和总VFA 含量均比空白对照组表现出显著提升(P<0.05)或极显著提升(P<0.01);酶B 组比酶A 组,乙酸和总VFA含量极显著提升(P<0.01),丙酸则呈现上升的趋势(0.05<P<0.1)。

表1 主效酶(Xyn)和辅效酶(AbfB、FAE)协同对断奶仔猪生长性能的影响
试验得出的结果表明:在含有阿拉伯木聚糖AX的纤维饲料原料中,添加AbfB、FAE能够与主效酶发挥协同作用,提高主链酶的降解效率,增加总还原糖的释放量,从而更充分地分解日粮中的抗营养成分,促进养分消化和吸收,提升断奶仔猪的日增重和日采食量,优化动物的肠道环境,降低料肉比和腹泻率(郑达文,2019)。
雷钊(2017)进行了肉鸡方面的试验,研究木聚糖酶与支链酶协作产生的产物作为益生元对肠道乳酸菌增殖的影响。
图4显示,图4a:未经酶解的麦麸表明光滑;图4b:木聚糖酶处理麦麸表面结构受到了破坏呈现典型的蜂窝状结构;图4c:额外添加阿拉伯呋喃糖苷酶;图4d阿魏酸酯酶都可以进一步破坏其表面结构。图4e:同时添加2 种支链酶,麦麸表面的蜂窝状结构最为明显。扫描电镜较为形象地证明了2 种支链酶与内切型木聚糖酶的协同效应。

图4 去淀粉麦麸经木聚糖酶、阿拉伯呋喃糖苷酶和阿魏酸酯酶酶解处理后的扫描电镜
添加木聚糖酶可以增加1~21 d和1~36 d肉鸡的平均日增重,并显著降低肉鸡的料肉比(P<0.05);额外添加阿拉伯呋喃糖苷酶和阿魏酸酯酶均可以进一步改善木聚糖酶的效果,提高肉鸡的生长性能,在肉鸡36 d 时,与单独添加木聚糖酶组相比,同时添加3 种酶制剂可以显著提高肉鸡的平均日增重(P<0.05),降低料肉比(P<0.05)(雷钊,2017)。由阿拉伯呋喃糖苷酶、阿魏酸酯酶和内切型木聚糖酶组成的特异性木聚糖降解酶,能有效地降解木聚糖,生成XOS。XOS 提高乳酸菌内编码ABC 转运蛋白和碳水化合物代谢中的关键酶基因的表达,促进乳酸菌的增殖,产生更多的丁酸,进而改善肉鸡的肠道健康和生产性能。
综上所述,郑达文(2019)和雷钊(2017)分别进行的猪鸡的试验,额外添加阿拉伯呋喃糖苷酶和阿魏酸酯酶这两种木聚糖的支链酶(侧链酶),比单纯使用木聚糖酶这种木聚糖主链酶效果更好,无论是还原糖、木糖、木寡糖释放,还是体外仿生消化效率,甚至动物生产性能及肠道微生物菌群和产物等方面都有差异和积极的效果。
上面讨论到的这种主链酶与侧链酶的相互配合、协同作用的存在,说明了Bhat等(2001)在讨论自然条件下复杂底物的酶作用存在“多酶复合体”,而他们同时也提出了是否其中各个单酶有相对效率之问,以此出发,在纤维类日粮中使用木聚糖酶及相关糖苷酶的试验基础上,我们提出了“主效酶”与“辅效酶”的概念,实际上,这种主次之分的酶制剂也可能存在于其他类成分,如蛋白与植物盐结合体,甚至脂类成分等。这里特别比较一下主效酶和辅效酶与常常提到的主链酶及侧链酶(或支链酶)的关系,后者的范围更窄,只是适合物料底物是以链状存在,其实除了化学组分差别影响酶发挥作用有别外,物理结构不同同样造成酶作用的变异。这两种情况体现在组合酶与配合酶的单酶关系。如果把动物日粮效果看成一个整体,复合酶这类在作用物理结构及化学成分关联度不高的酶,同样也存在一种主次之别,存在主效酶与辅效酶的区别。这种单酶之间关系与效率的区分,为饲料酶制剂的设计与配制的科学基础,饲料酶使用的选择与性价比的考虑都有一定的意义,可以进一步深化和细化饲料酶的应用。
