非生长时期6种植物枝条的抗折力学特性
2019-10-11格日乐郝需婷苏日娜崔天民
格日乐, 郝需婷, 苏日娜, 黄 静, 崔天民
(内蒙古农业大学 沙漠治理学院, 内蒙古 呼和浩特 010019)
在治理土壤风力侵蚀的过程中,植被通过遮盖地面、分化风力以及拦截输沙等办法有效地减弱风速、削减风对地面土粒的分离和搬运而起作用。关于植物防风抗蚀的研究,国外学者Bressolier和Thomas[1]在法国大西洋沿岸沙丘区的研究结果表明,在植被的诸多特点中,对风沙运动植被盖度作用最大,然后是植被的高度和宽度。刘艳萍等人[2]通过风洞试验研究了荒漠草原区植被对土壤风蚀的影响,证明了风速越大植被对地表的保护作用越明显,相关分析表明,不同风速下风蚀量与植被盖度之间呈幂函数相关。孙艳萍,张晓萍,刘建祥等人[3]研究得出,风蚀率随植被盖度的增加呈指数函数减少。不同植物防治风蚀的性能是不同的,赵彩霞等人[4]研究表明,在干旱、半干旱地区灌木的防风蚀作用最大,其次分别是多年生牧草、林木、作物、一年生牧草。Lee和Soliman总结了大量试验观测结果后指出,植株的形状是影响植被覆盖防护效应的重要因素,植被类型的不同可能造成防护效应的显著差异[5]。综上所述,关于植被防风抗蚀的研究,主要集中在植被盖度、植被类型以及植株形状(高度、宽度)等方面。内蒙古中西部地区冬春季正值大风季节,当大风过境时,植物体承受强大的风力作用。风力作用在植物体上,被枝茎阻挡牵引等消耗一部分能量,越过植物体后力量减小,在植被防风抗蚀中地上部分枝条力学特性亦发挥重要的作用。而现有的研究,主要是针对植物生长季节根系[6-8]和小麦(Triticumaestivum)[9-10]、玉米(Zeamays)[11]、高粱(Sorghumbicolor)[12]、水稻(Oryzasativa)[13-16]等特定茎秆作物的抗倒伏性,及针对植物在生长季鲜枝力学特性的研究[17-18],而关于植物在非生长时期的冬春大风季节枝条力学特性的研究鲜见报道。为此,本文选取分布于内蒙古自治区中西部的6种典型的防风固沙植物(沙柳、柠条、白沙蒿、杨柴、柽柳、沙棘),以非生长时期的冬春交替大风季节植物枝条为对象,通过研究非生长时期枝条抗折力和抗折强度及其影响因子,分析直径、长度和植物种对枝条抗折力、抗折强度的影响,揭示其种间差异性以及抗折力学特性的共同规律,从而为该区域防风阻沙植物种的选择及其混交配置提供生物力学方面的科学参考依据。
1 材料及采样地概况
沙柳、柠条、白沙蒿、杨柴和柽柳采样地位于库布齐沙漠西北部,库布齐沙漠地处鄂尔多斯高原北部与河套平原的交接地带,地理位置为107°—111°30′E,39°30′—39°15′N,呈东西带状分布,长约400 km,枝条采集样地位于库布齐沙漠西北端的固定沙地,采样地中心位置为39°26.89′N,107°23.85′E。沙棘采样地位于鄂尔多斯市准格尔旗东南部黄土丘陵沟壑区。
沙柳(Salixcheilophila)为杨柳科(Salicaceae)柳属(Salix),是典型的旱生落叶灌木或小乔木,沙柳属沙生阳性植物,抗逆性强,较耐旱,抗风沙、水涝、牛羊啃、刀斧砍,耐一定盐碱,耐严寒和酷热,喜适度沙压,根系发达,萌蘖力强。沙柳的根系最长可延伸至100 m,一株沙柳就可将周围流动的沙漠牢牢固住,因此有很好的固沙作用,大面积的沙柳分布具有改善小气候作用,可有效遏制频发的沙尘暴天气,降低地表温度和水分蒸发量,减轻霜冻和干旱危害,是地下资源开发地区良好的生态屏障。此外,沙柳是制作生物沙障的主要树种,通过栽植沙柳在沙面上设置障碍,以控制风沙流动的方向、速度、结构来改变风蚀积沙状况,达到防风阻沙、改变风的作用力及地貌状况等目的,是固定流沙、稳定地表首选的生物措施[19]。柠条(CaraganaKorshinskii)是豆科(Leguminosae)锦鸡儿属(Caragana)多年生灌木,耐旱、耐寒、耐高温、耐贫瘠、耐盐碱性,不怕沙埋,是植物王国中的“铁人”、“铁汉子”。根系发达,而且其蒸发量很低,能够最大程度的保水、吸收地下深处的营养物质与水分。柠条苗木比较容易培育,而且能够顺利在沙地中成活,成活以后,对于沙地能够起到固沙固氮的作用,同时在柠条成活以后,可以适当引进其他树种,这样柠条就起到在治沙造林中的先锋作用,为其他树种成活铺路[20]。白沙蒿(Artemisiasphaerocephala)又称籽蒿,属于菊科(Asteraceae)蒿属(Artemisia)典型的沙生半灌木,生于流动与半流动以及半固定沙丘上,是固沙的先锋植物,具有耐沙埋、抗风蚀、耐贫瘠、抗旱、易于繁殖等特性。具有较强的生态适应性,是重要的防风固沙植物[21]。沙棘(Hippophaerhamnoides)属于胡颓子科(Elaeagnaceae)沙棘属(Hippophae)落叶灌木或小乔木。主要分布于我国华北、西北和东北地区,适应性强,抗旱耐涝。沙棘有发达的根系,其根系可深达地面高度的5倍,须根横向生长可达5 m多,每株沙棘固沙保土面积可达70~80 m2,固沙能力之强,是其他乔木、灌木无法比拟的。不仅如此,沙棘还耐干旱、耐瘠薄,生存能力极强,栽培沙棘树苗时,每株只需约1 kg的水即可成活,成活后还可通过种子以及裸露的根系繁殖,能迅速扩大林木面积。所以沙棘是治理我国华北、西北地区风沙的理想植物之一[22]。杨柴(Hedysarumfruticosum)属于蝶形豆科(Leguminosae)岩黄蓍属(Hedysarum)多年生、沙生落叶半灌木。杨柴地下根茎的生长纵横交错,形成一个网带结构的水平空间,这种网带结构的存在,起到了很好的固定沙丘的作用。而且杨柴能够产生适应沙埋的生理机制,使杨柴能够在适度沙埋后生长更旺盛。杨柴具有耐寒、抗旱、耐瘠薄,抗风蚀、抗沙埋和易栽植,生长迅速,竞争力强的优良特性,是优良的防风固沙植物。杨柴是我国北方的毛乌素沙地、库布齐沙漠和浑善达克沙地的治理中被作为主要植物种进行了大面积种植[23]。柽柳(Tamarixchinensis)属于柽柳科(Tamaricaceae)柽柳属(Tamarix)灌木或小乔木。柽柳属于泌盐盐生植物,具有很强的耐盐碱能力,是盐碱地种植的首选灌木。柽柳的种植可以在一定程度上改变盐碱地的土壤性状,增加土壤的植被盖度,使盐分发生转移,增加土壤有机质的含量,改善土壤肥力,对盐碱地起到改良作用,并显著提高盐碱地生态与环境功能[24]。柽柳也是最适应干旱沙漠的树种之一,根系很长,可以吸到深层的地下水,枝条顽强不怕沙埋,是防风固沙的优良树种之一。
2 研究方法
2.1 标准株的确定与枝条的采集
于2018年3月上旬,在采样地选择柠条、沙棘、沙柳、白沙蒿、杨柴和柽柳6种植物样地,在每种植物的样地内选择30株植株,将每一株植物的冠幅、地径和高度重复测量3次。再将30株植株的高度、冠幅和地径取平均值来作为衡量该植物种的综合指标。在每个样地内尽量选取与综合指标相近的植株作为标准株[25]。选择标准株3株,在标准株的植株上尽量选择无病虫害和人为破坏的枝条进行采集。将采集的枝条带回试验室内,尽快完成试验。
2.2 试样制备
选择枝条较为通直、直径较为均匀的枝条。根据前期对6种植物枝条数量特征的研究结果[25],试样枝条直径选取0.5~2.5 mm范围,以0.5 mm为一个间隔进行分级,选取60,80和100 mm共3个长度梯度。当试样长度为60 mm时非生长时期从枝条一侧开始每15 mm作1个标记一共做5个标记(D1—D5)。在试验枝条两端标记点的一侧贴上编有序号的标签,用游标卡尺从标签一侧点开始依次量取5个标记点处直径,正交方向上各测量一次直径,然后取5个点的直径平均值作为该试验枝直径D。将制备好的试验枝条立即进行试验。枝长80和100 mm下的标记及测量方法与60 mm时一致。采用两端固定式折断法进行试验研究。试验仪器如图1所示,包括YG(B)026H-250型织物强力机、自制夹具以及电子游标卡尺(精度0.01 mm)。试验开始前调节仪器以及联机软件的相关参数后,用自制夹具将试验枝条两端的标记点夹紧,在500 mm/min的加载速度下开始试验,每一次压头折断枝条施力点时试验结束。试验结束的同时,联机软件会相应的自动生成本次试验枝条抗折力值(F),将此时的数据在试验记录本上记录,最后根据所记录的数据结合抗折力计算公式计算抗折强度,计算公式为:
式中:σ——抗折强度(MPa);F——抗折力(N);D——直径(mm)。

图1 抗折试验方法示意图
3 结果与分析
3.1 非生长时期6种植物枝条抗折力及其影响因子
运用SPSS软件对6种植物枝条在60 mm长度下种内不同径级抗折力进行差异性检验结果显示,在α=0.05的显著水平下,除了沙柳非生长时期枝条抗折力在1~1.5 mm和1.5~2 mm径级之间无显著差异以外其他径级间均呈显著差异,另外5种植物枝条抗折力在直径0.5~1 mm,1~1.5 mm,1.5~2 mm和2~2.5 mm径级之间均存在显著性差异,表明直径因素对6种植物枝条抗折力大小有显著的影响,抗折力随着直径的增加而增大(表1)。

表1 非生长时期6种植物枝条抗折力在不同径级种内差异性检验结果 N
注:数据为均值±标准差;相同植物种不同径级间字母不同则表示抗折力差异性显著,字母相同或包含相同字母则表示差异性不显著(p<0.05)。下同。
图2为6种植物枝条在60 mm长度下抗折力随直径变化图。由图2可知,6种植物枝条抗折力均随直径的增加而增大。沙柳枝条直径与抗折力呈对数函数正相关关系、沙棘呈幂函数正相关关系,其他4种植物非生长时期枝条抗折力与直径的关系均呈多项式函数正相关关系(表2)。这与郑永刚[7]选取柠条、沙棘、沙柳和白沙蒿的直根抗折力随直径增大而增加的结果一致,只是拟合函数不同,分析原因是因为供试植物均属于干旱半干旱区灌木,而灌木枝条和根系均属于植物材料,材料性质相同的缘故,拟合函数不同则是因为植物种不同则生物学特性不同,关于上述结果还需要结合材料微观结构和化学组分等进行进一步研究。
表3为运用SPSS软件对6种植物枝条在60,80和100 mm长度下相同径级范围(0.5~2.5 mm)内种间抗折力的差异性检验结果表。由表3可知,在60 mm枝长下,相同径级不同植物种间,在α=0.05的显著水平下,柠条抗折力与沙棘、柽柳、沙柳、杨柴和白沙蒿5种植物枝条抗折力均存在显著差异性,而沙棘、柽柳和沙柳3种植物枝条抗折力间无显著差异性,杨柴和白沙蒿枝条抗折力间无显著差异性。6种植物枝条抗折力大小则表现为:柠条(51.700 N)>沙棘(39.706 N)>柽柳(37.530 N)>沙柳(36.156 N)>杨柴(29.553 N)>白沙蒿(27.147 N)。

图2 非生长时期6种植物枝条抗折力随直径变化表2 6种植物枝条抗折力与直径的拟合函数

植物种拟合函数决定系数R2柠条y=25.814x2-27.631x+26.8520.9752沙柳y=19.368ln(x)+30.5090.9333杨柴y=13.864x2-13.494x+16.9980.987沙棘y=20.12x1.4620.9611柽柳y=34.096x2-49.551x+26.1560.9771白沙蒿y=19.817x2-32.919x+26.60.9741

表3 非生长时期6种植物枝条在60 mm相同径级下种间抗折力差异性检验结果 N
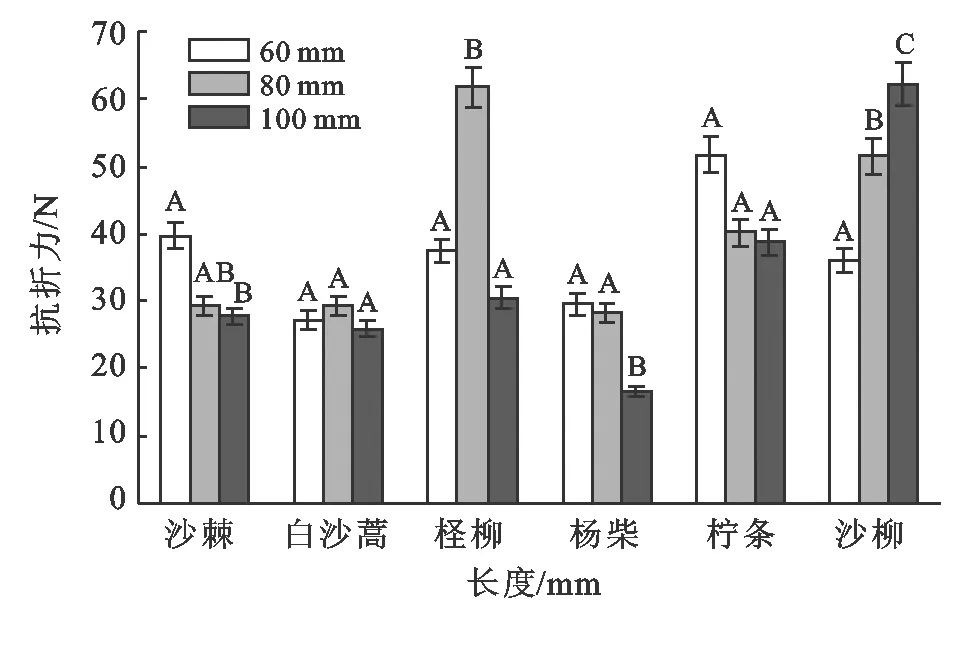
在80 mm枝长下,相同径级范围(0.5~2.5 mm)内在α=0.05的显著水平下,白沙蒿、沙棘和杨柴3种植物枝条抗折力间无显著差异性以外其他均存在显著差异性;6种植物枝条抗折力大小依次为:柽柳(61.777 N)>沙柳(51.440 N)>柠条(40.180 N)>白沙蒿(29.370 N)>沙棘(29.270 N)>杨柴(28.401 N)。当枝条在100 mm长度下,在α=0.05的显著水平下,沙柳、杨柴枝条抗折力与其他4种植物柠条、柽柳、沙棘、白沙蒿存在显著差异性,而柠条、柽柳、沙棘、白沙蒿4种植物枝条抗折力间差异性不显著;6植物枝条抗折力大小表现为:沙柳(62.237 N)>柠条(38.889 N)>柽柳(30.575 N)>沙棘(27.768 N)>白沙蒿(25.875 N)>杨柴(16.590 N)。从图3中可以看出,在α=0.05显著水平下,白沙蒿和柠条枝条抗折力随着枝条长度的增加无显著性差异,表明枝长对2种植物枝条抗折力变化无影响。沙柳则相反,随着枝条长度的增加抗折力呈增大趋势,表明枝长对沙柳枝条抗折力有明显影响。其他3种植物沙棘、柽柳和杨柴枝条抗折力随着枝条长度的增加无明显变化规律。上述结果表明植物种不同则枝长对枝条抗折力的影响亦有差异,关于这点还需要结合枝条微观结构及其纤维素、半纤维素、木质素等化学组分进行进一步研究。
3.2 非生长时期6种植物枝条抗折强度及其影响因子
运用SPSS软件对6种植物枝条在60 mm长度下种内不同径级间抗折强度进行差异性检验结果显示,在α=0.05的显著水平下白沙蒿枝条抗折强度在直径1.5~2.0 mm,2~2.5 mm之间无显著差异以外,其他植物种枝条径级之间均存在显著差异性,表明直径因素对枝条抗折强度有显著影响,抗折强度随直径的增加而减小(表4)。
注:图中包含相同字母则表示在0.05水平下差异性不显著,字母不同则表示差异性显著(LSD多重比较结果)。下同。
图3 非生长时期6种植物枝条抗折力随枝条长度变化

表4 非生长时期6种植物枝条种内不同径级间抗折强度差异性检验结果 MPa
图4为6种植物枝条在枝长60 mm时抗折强度随直径变化图。由图4可知,在非生长时期6种植物枝条抗折强度均随直径的增加而减小。6种植物枝条抗折强度与直径的关系均呈幂函数负相关关系(表5)。这与郑永刚[7]关于柠条、沙棘、沙柳和白沙蒿直根的研究结果一致,只是变化函数不同,其原因与枝条抗折力的结果分析一致。

图4 非生长时期6种植物枝条抗折强度随直径变化
表6是运用SPSS软件对6种植物枝条在枝长60,80和100 mm长度下相同径级范围(0.5~2.5 mm)内种间抗折强度的差异性检验结果。由表6可知,在α=0.05的显著水平下,当枝长为60 mm时,柠条抗折强度与沙柳抗折强度无显著性差异,杨柴、沙棘、柽柳和白沙蒿抗折强度间无显著差异性;6种植物枝条抗折强度大小依次为:柠条(33.510 MPa)>沙柳(30.314 MPa)>杨柴(23.783 MPa)>沙棘(22.500 MPa)>柽柳(20.044 MPa)>白沙蒿(16.613 MPa)。

表5 6种植物物枝条抗折强度与直径的拟合函数
当枝条长度在80 mm时,在α=0.05的显著水平下,柽柳与沙柳枝条抗折强度差异性不显著,白沙蒿和柠条枝条抗折强度无显著性差异,二者又与沙柳枝条抗折强度差异性不显著;沙棘和杨柴枝条抗折强度无显著性差异。在80 mm枝条长度下,6种植物枝条抗折强度大小依次表现为:柽柳30.821 MPa,沙柳27.122 MPa,白沙蒿23.013 MPa,柠条22.932 MPa,沙棘16.778 MPa和杨柴16.047 MPa。当枝条长度在100 mm时,在α=0.05的显著水平下,沙柳抗折强度与其余5种植物均存在显著性差异;白沙蒿和柠条枝条抗折强度差异性不显著,沙棘、柽柳和杨柴枝条抗折强度间无显著性差异,柠条又与沙棘、柽柳和杨柴3种植物枝条抗折强度间差异性不显著。6种植物枝条在长度为100 mm时,种间抗折强度大小表现为沙柳最大为33.156 MPa,其次为白沙蒿20.030 MPa,柠条18.837 MPa,沙棘14.262 MPa,柽柳12.718 MPa,抗折强度最小为杨柴9.620 MPa。
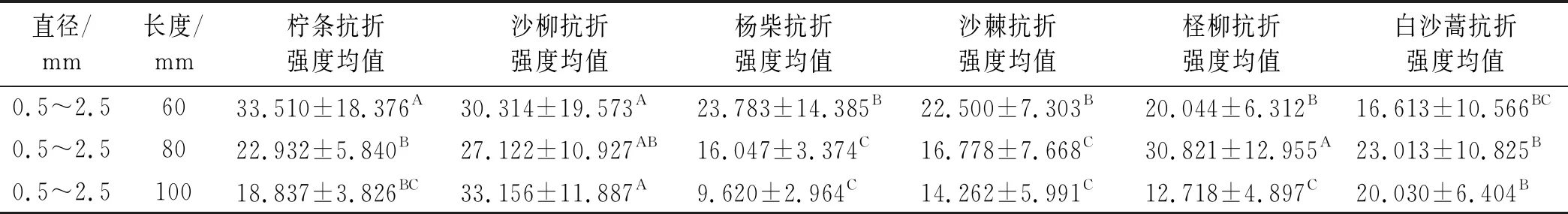
表6 6种植物非生长时期枝条在60 mm相同径级下种间抗折强度差异性检验结果 MPa
图5为6种植物枝条抗折强度随枝长变化图。从图5可以看出,在α=0.05显著水平下,沙柳枝条抗折强度随枝长的增加无显著差异性,表明枝长对其枝条抗折强度无影响,而其他5种植物枝条抗折强度随枝条长度的增加变化不一致,其中杨柴、柠条和沙棘总体随长度的增加枝条抗折强度呈下降趋势,而白沙蒿和柽柳则呈现先增加后减小的变化。
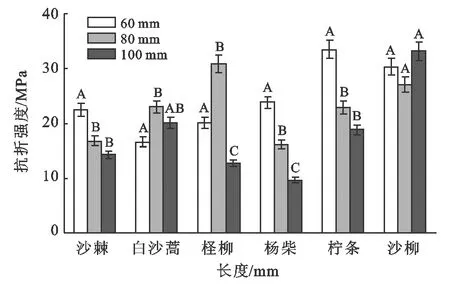
图5 非生长时期6种植物枝条抗折强度随枝长变化
4 讨 论
6种植物在非生长时期枝条抗折力随着直径的增加而增大,抗折强度随直径的增加均呈幂函数减小,这与郑永刚[7]对柠条、沙棘、沙柳和白沙蒿直根的研究结果一致,只是随枝条直径的增加,抗折力增大或抗折强度减小的变化函数有所不同。这可能是因为上述对比分析的植物均属于干旱半干旱区灌木植物,植物材料性质相同的缘故,变化函数不同是由于植物种不同则生物学特性不同,其内在原因还需要结合材料微观结构和化学组分等进行进一步研究。关于枝条或根系长度对植物枝条或根系抗折力学特性研究的文献很少,薛冬梅[17]在3种植物枝条生物力学特性的研究结果中表明枝长对植物枝条的抗拉强度无显著影响,抗弯强度则随枝条长度的增加而增大。本文通过对6种植物非生长时期枝条抗折力和抗折强度随着枝条长度的增加,研究结论显示植物种不同则枝长对枝条抗折力和抗折强度的影响亦有差异,即有些植物种表现为枝长因子对其枝条抗折力学强度指标无明显影响,而另外一些植物种则表现为枝长因子对其枝条抗折力学强度指标的变化有明显影响,究其内在原因还需要结合不同植物种枝条微观结构及其纤维素、半纤维素、木质素等进行进一步研究[26-27]。同时这也是否从另外一个方面证明了生态修复中只有实现多树种结合的修复方式才是最优水土保持功能实现的模式。
5 结 论
(1) 6种植物枝条在非生长时期抗折力、抗折强度在0.5~2.5 mm测试径级范围,随着直径的增加抗折力呈现增大趋势,而抗折强度则呈现减小的趋势。其中沙柳枝条抗折力呈对数函数增长、沙棘枝条抗折力呈幂函数增长,其他4种植物枝条抗折力随着直径的增加均呈多项式函数增长;6种植物枝条抗折强度随直径的增加均呈幂函数减小。
(2) 在测试径级0.5~2.5 mm范围内枝长在60 mm枝条抗折力、抗折强度最大为柠条,其值分别为51.700 N和33.510 MPa,枝长在80 mm时种间抗折力、抗折强度最大为柽柳,其值分别为61.777 N和30.821 MPa,枝长在100 mm时种间抗折力、抗折强度最大为沙柳,其值分别为62.237 N和33.156 MPa。表明枝长不同则植物种间枝条抗折力和抗折强度值有差异。证明生态修复中只有实现多树种结合的修复方式才是最优水土保持功能实现的模式。
(3) 6种植物在非生长时期枝条抗折力和抗折强度随着枝条长度的增加其变化特点表现不一致,关于这点还需要结合不同植物种枝条微观结构及其纤维素、半纤维素、木质素等化学组分进行进一步研究。
