宁夏云雾山典型草原物种功能性状和群落功能多样性对氮素添加的响应
2019-09-17李维军
李维军

摘要:基于植物功能性状的群落功能多样性是影响生态系统功能的主要因素。研究群落水平植物功能性状及其功能多样性随氮素添加浓度的变化规律,对认识不同氮素添加浓度下植物群落的构建以及指导退化草地恢复重建具有重要意义。以宁夏回族自治区云雾山国家自然保护区不同施肥梯度的典型草原为研究对象,比较了植物功能性状(株高、叶氮含量、比叶面积)和3种功能多样性指标(功能丰富度、功能均匀度、功能离散度)随施肥梯度的变化规律。结果表明,群落地上生物量随氮素添加浓度的增加逐渐增加。主成分分析表明,叶片氮含量、植株最大高度和比叶面积可作为氮素添加后的指示性状;随着氮素水平的提高,最大植株高度的群落加权性状值(群落中每个物种的功能性状值与其相对多度相乘后的总和)持续增加,比叶面积的群落加权性状值变化较小,叶片氮含量群落加权性状值表现为先增加后降低的趋势。物种多样性(物种丰富度指数和多样性指数)随氮素添加浓度的增加呈现为降低趋势;而功能多样性保持相对稳定,不同氮素水平间差异不显著。由物种功能性状决定的物种多样性丧失过程可能比群落功能多样性更能直接地反映出氮素添加后群落结构的变化。研究结果对宁夏以及黄土高原同类地区的植被恢复重建的物种选择及植被布局规划具有重要的实践意义。
关键词:氮添加;功能性状;功能丰富度;功能均匀度;功能离散度
中图分类号: S181 文献标志码: A 文章编号:1002-1302(2019)07-0282-05
我国的各类草地有4亿hm2,占国土面积的41.7%,其中大部分分布在北方,是我国主要的草牧业生产基地和重要的生态安全屏障。然而,受全球气候变化等自然因素和长期过度放牧、乱垦乱挖等人为因素的综合影响,我国草地约90%以上的面积出现不同程度的退化,这导致了生产和生态功能严重下降[1]。我国政府从20世纪末开始,针对草地退化、畜牧业发展效益低下等难题,制定并实施了一系列工程和政策措施,目前草地严重退化现象已初步得到遏制,但“局部改善,整体退化”的态势并未得到有效改观。
黄土区的草地面积为2.32×107 hm2,占其土地总面积的33%[2],其中以本氏针茅(Stipa bungeana)为建群种的典型草原为主体,占黄土高原草地面积的60%,这在维持区域生态平衡、调节气候以及保持水土等方面占有重要地位[3]。深入研究本氏针茅草原恢复管理过程中生物多样性变化及生态系统功能,对于改善脆弱的黄土区生态系统功能及维护、建设西北地区生态屏障具有重要科学意义。
以往国内外生态学者在描述群落的组成和结构与生态系统功能的关系时,常采用物种丰富度或物种多样性指标,并将他们等同于生物多样性[4-5],往往忽略功能多样性(functional diversity)这个重要的生物多样性指标。功能多样性是指特定生态系统中物种功能性状的数值和范围,也称为功能性状多样性(functional trait diversity),它强调群落中物种功能的差异[6]。相对于基于物种结构特征的物种多样性,基于功能性状的群落功能多样性才是影响生态系统功能和过程的关键所在[7]。我国植物功能性状的尝试性工作开始于我国东北样带草原植物性状与降水梯度和资源利用等的关系[8]以及青藏高原植物生理生态学功能性状的研究[9],沿环境梯度的植物功能性状研究将对我国全球变化领域的研究有所贡献,但基于植物功能多样性与生态系统功能的研究刚刚起步[10-12],且在恢复草地管理利用(施肥、刈割、放牧等)过程中的研究更鲜有报道。
鉴于此,本试验以宁夏回族自治区云雾山国家自然保护区典型草原为研究对象,以植物的功能性状为切入点,试图揭示草地不同氮素添加浓度下物种功能性状和群落功能多样性变化特征,以期逐步建立、完善适合宁夏及黄土高原同类地区天然草地功能性状的体系及其研究方法,为黄土高原天然草地的后续合理利用以及我国生态文明建设和美丽中国战略目标的实现提供理论依据。
1 材料与方法
1.1 研究地概况
研究地点位于宁夏回族自治区固原市东北部的云雾山国家级草地自然保护区(106°21′~106°27′E,36°10′~36°17′N),海拔高度1 800~2 100 m。该区域气候属与中温带半干旱区,年平均气温5 ℃,最冷月为1月(平均气温-14 ℃),最热月为7月(22~25 ℃),≥0 ℃的积温为2 370~2 882 ℃,年日照时数为2 500 h,太阳辐射总量 125 kcal/cm2,年降水量400~450 mm,降水季节分配不均,全年65%~75%的降水集中在7—9月。蒸发量1 330~1 640 mm,干燥度1.5~2.1,无霜期112~140 d。灾害性天气主要有干旱、暴雨、霜冻、冰雹、干热风等。
保护区植被类型属温带典型草原,其建群种和优势种以本氏针茅(Stipa bungeana)、百里香(Thymus mongolicus)、白莲蒿(Artemisia sacrorum)、大针茅(Stipa grandis)、冷蒿(Artemisia frigida)群落为主,伴生种类型以猪毛蒿(Artemisia scoparis)、百里香(Thymus mongolicus)、星毛委陵菜(Comarum acaulis)群落为主,其中丛生禾本科植物本氏针茅在该区分布范围最广。
1.2 试验设计
试验选取长期封育的典型草原为研究对象,采取隨机区组设计进行长期氮素添加试验[13]。以尿素作为肥料,设置5个氮素添加水平:CK,0 g/(m2·年);N5,5 g/(m2·年);N10,10 g/(m2·年);N20,20 g/(m2·年);N40,40 g/(m2·年),每个水平重复8次,即每块样地上设置40个6 m×10 m小区,其中一半(3 m×10 m)用于群落植被调查,一半(3 m×10 m)用于植物个体采样,各区间有2 m缓冲带,于生长季初即每年4月底进行施肥。
1.3 群落调查和植物功能性状测定
在植物生长旺盛期间(2017年8月下旬)于每个4 m×10 m 的群落调查小区内中随机选取1个1 m×1 m的样方,用群落学调查方法测定群落中的物种组成,包括物种数目、植株高度、各物种多度及盖度、用收获法测定地上总生物量,表征生态系统功能,并按物种分类,带回实验室80 ℃烘干至恒重。
在每个重复小区内,每个物种采集3株成熟、完整、无病虫害个体的地上部分,装入塑封袋带回,在室内测量其株高;在每个植株的中部各选取2张成熟、展开、无病虫害的叶子,进行植物叶性状的测量。用电子秤称取叶片鲜质量,用CI-202叶面积仪扫描叶面积;然后装入纸袋中,在80 ℃温度下烘48 h,称其叶片干质量。
1.4 数据分析
利用Microsoft Excel 2007和R 3.3.1整理计算、分析数据及作图。采用One-way ANOVA和Tukey法进行显著性检验和多重比较,利用Pearson法进行相关性检验。群落功能多样性各指标均通过F-diversity软件计算获取。运用主成分分析(PCA)从众多植物功能性状中筛选出主要功能性状因子。
2 结果与分析
2.1 群落地上生物量对不同氮素添加水平的响应
地上生物量是生态系统功能最集中的体现。一般而言,养分添加能导致群落地上生物量增大,物种多样性减少。由图1可知,随着氮素浓度的增加,地上生物量逐渐增大。
2.2 植物功能性状的变化特征
采用主成分分析方法在多个植物功能性状中筛选出最为敏感和重要的功能性状,结果见表1。在所选功能性状中可划分3个主成分,最高累计方差为88.25%。根据主成分分析的结果和植物生态对策[17-18],选取叶片氮含量、最大植株高度、比叶面积来表征氮素添加后植物功能性状的变化特征。
由表2和图2可知,在不同氮素添加浓度下,3个植物功能性状间相互独立,且变化趋势差异明显。植物最大高度的群落加权性状值(群落中每个物种的功能性状值与其相对多度相乘后的总和)随着氮素添加浓度的增大表现为持续增加,这是因为添加氮素使得物种从地下竞争(土壤资源)转化为地上竞争(光资源),相对较高的物种(如白莲蒿和甘青针茅、大针茅)成为优势种,导致一些低矮或较小的植物(如二裂委陵菜、星毛委陵菜)数量减少。随着氮素添加浓度的增大,比叶面积的群落加权性状值变化较小,仅N5和N10差异显著,其他处理间差异并不显著,这与物种的个体差异有关。叶片氮含量的群落加权性状值随着氮素水平的提高表现为先增加后降低的趋势。因此,最大植株高度和叶片氮含量的加权性状值的变化特征表明,一些对氮素敏感的物种(快速投资-收益型)能够利用外源氮素迅速生长、代谢从而在群落中占据优势地位,而一些对氮素不敏感的物种(慢投资-保守型)则表现为生长缓慢,在群落中处于劣势地位,甚至被竞争排除,氮素添加快速投资-收益型的物种优势凸显。
2.3 物种多样性和功能多样性的变化特征
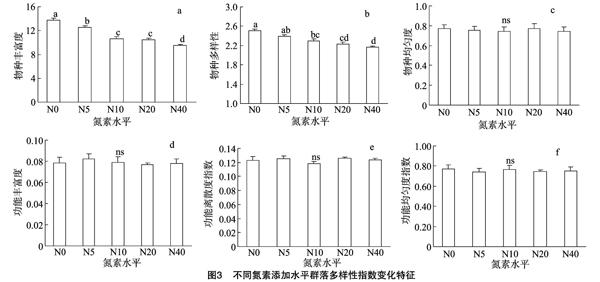
从表3可以看出,物种多样性指数和物种丰富度指数有一定的相关性,但功能多样性指标(功能离散度、功能均匀度和功能丰富度)和物种多样性指标(物种多样性、物种丰富度和物种均匀度)间的相关性不显著,且相关性系数的绝对值较小;说明功能多样性和物种多样性之间可能是相互独立的。在不同氮素添加水平下,功能多样性变化与物种多样性的变化趋势不尽相同。从图3可看出,物种多样性各指标随着氮素添加水平的增加变化趋势也不相同。图3-a、3-b显示,随着氮素添加水平的增加,物种丰富度指数和物种多样性指数均表现为逐渐减小的趋势。与对照样地N0相比,N5、N10、N20和N40的物种丰富度指数分别降低了9.09%、22.73%、23.64%、30.91%;其中,N10和N20差异不显著,其他氮素添加水平间均差异显著;物种多样性指数则分别减少了4.63%、8.45%、10.96%、13.84%;图3-c显示氮素添加后,物种均匀度指数并未发生显著性变化。图3-d、3-e、3-f 表明,功能丰富度指数、功能离散度指数和功能均匀度指数在不同氮素添加水平下相对稳定,差异不显著。
3 讨论与结论
3.1 群落地上生物量对氮素添加的响应
草地群落生物量可反映植被的功能,其大小受降水、温度和土壤养分等非生物因素和物种之间的竞争、正相互作用等生物因素的影响。本试验结果表明,氮素添加明显增加了地下土壤有效资源,进而缓解了植物生长的養分限制,促进了植物地上部分的生长,从而明显增加了地上生物量,这与多数研
究结果[19-20]相一致;而施氮浓度越高,地上生物量的增加量越多,这与内蒙古的草甸草原、典型草原、荒漠草原以及青藏高原的高寒草甸研究结果[20-22]相似。
3.2 植物功能性状
植物功能性状能够反映植物组织结构和功能的平衡[23],不同植物功能性状对氮素添加响应因自身系统发育的不同存在较大差异。肖迪等研究认为当环境改变时,植物会改变其生长策略,重新调整资源分配,从而保证物种在整个群落中的地位和群落结构的稳定性[24]。万宏伟等通过对内蒙古羊草群落常见优势种的比叶面积、叶片含氮量等叶片功能特性对氮素响应的研究证实了氮素添加改变了群落的光合有效辐射[25]。杨浩等研究发现,氮素添加主要改变了糙隐子草的叶形态和植物的一些生理性状,而不是改变植物整株的性状[26]。为避免物种不同引起的响应差异,本试验综合了10个所选物种的功能性状,通过主成分分析筛选出敏感性状来分析群落功能多样性的变化规律。结果表明,最大植株高度、叶片氮含量、比叶面积这3个植物功能性状可以作为评价不同氮素添加水平的响应性状。最大植株高度的群落加权性状值随氮素添加呈增加的趋势,同时叶片氮含量的群落加权性状值随氮素添加呈先增加后降低的趋势,表明高度较低、生长较慢的物种最终被高度较高、生长迅速的物种所替代。氮添加可使植物在性状空间中的位置向由“慢投资-保守型”向“快速投资-收益型”发生转变。
3.3 物种多样性和功能多样性
群落多样性对氮素添加的的响应与其所处环境条件与土壤养分条件密切相关。有研究表明,氮素添加改善了退化草地的土壤状况,增加了物种多样性[27];也有研究认为,氮素添加对物种多样性并无显著影响[28];还有研究显示,物种多样性随着氮素添加浓度的增加呈现先增加后降低的趋势[29]。但绝大部分研究说明,氮素添加导致物种丰富度和物种多样性指数的下降[30-32],其原因可能是(1)氮素添加提高了养分有效性,促使植物由地下对养分资源的竞争转向地上光资源的竞争,一些低矮或者植株个体较小物种在竞争中处于劣势,甚至被竞争排除掉,从而降低了物种多样性[33];(2)氮素添加有可能增加地表凋落物数量,凋落物的遮光和阻隔作用可能抑制了一些物种的萌发或者幼苗的存活,进而降低物种多样性[34];(3)氮素添加(尤其是长期氮素添加或者氮素添加浓度过高)会引起土壤pH值降低或者产生铵中毒,一些不耐酸性和铵基毒性较弱的物种的生长受到抑制,甚至从群落中消失,从而降低物种多样性[35]。本试验表明,随着氮素添加浓度的增加,物种丰富度和多样性指数表现为降低的趋势。
氮素添加后群落功能多样性变化可由2个互为相反的机制决定,一是由于施肥后竞争排除和一些物种的消失可能引起功能多样性的降低[36];二是施肥后现存物种的生态位分化可能促进功能多样性的进一步增加[16];在此情况下,由施肥引起的竞争有可能会限制物种的极限相似性,进而使物种性状逐渐产生差异[37]。氮素添加后群落功能多样性的变化最终取决于这2种机制的动态平衡。在本试验中,功能多样性并未呈现显著的增加或降低趋势,说明功能多样性指数对氮素添加并不敏感,这与Li等在青藏高原高寒草甸的研究结果[22]类似,可能是因为这2个互为对立的生态过程相互制约,一是氮素添加导致劣势物种被竞争淘汰,使得氮素添加群落的物种性状所占空间低于氮素未添加的群落[31];二是随着氮素的添加物种的功能性状呈不对称变化(如优势种的高度变化显著高于稀有物种),促进了氮素添加群落的生态位分化,补偿甚至抵消了物种消失引起的多样性的降低[38]。
综上所述,由物种功能性状决定的物种多样性的丧失过程可能比群落功能多样性更能直接反映氮素添加后群落性状结构的变化。本试验研究结果可为黄土高原天然草原的保护、合理利用以及草食畜牧业的发展具有重要实践意义。
参考文献:
[1]候向阳,尹燕亭,王婷婷. 北方草原牧户心理载畜率与草畜平衡生态管理途径[J]. 生态学报,2015,35(24):8036-8045.
[2]齐玉春,董云社,耿元波,等. 我国草地生态系统碳循环研究进展[J]. 地理科学进展,2003,22(4):342-352.
[3]程积民,程 杰,杨晓梅. 黄土高原草地植被与土壤固碳量研究[J]. 自然资源学报,2011,26(3):401-411.
[4]Adler P B,Seabloom E W,Borer E T,et al. Productivity is a poor predictor of plant species richness[J]. Science,2011,333(6050):1750-1753.
[5]王海东,张璐璐,朱志红. 刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制[J]. 植物生态学报,2013,37(4):279-295.
[6]Petchey O L,Gaston K J. Functional diversity:back to basics and looking forward[J]. Ecology Letters,2006,9(6):741-758.
[7]Butterfield B J, Suding K N. Single-trait functional indices outperform multi-trait indices in linking environmental gradients and ecosystem services in complex landscape[J]. Journal of Ecology,2013,101(1):9-17.
[8]Wang G H,Ni J. Responses of plant functional types to an environmental gradient on the Northeast China Transect[J]. Ecological Research,2005,20(5):563-572.
[9]He J S,Wang Z H,Wang X P,et al. A test of the generality of leaf trait relationships on the Tibetan Plateau[J]. New Phytologist,2006,170(4):835-848.
[10]孟婷婷,倪 健,王國宏. 植物功能性状与环境和生态系统功能[J]. 植物生态学报,2007,31(1):150-165.
[11]孔彬彬,卫欣华,杜家丽,等. 刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响[J]. 植物生态学报,2016,40(3):187-199.
[12]石明明,牛得草,王 莹,等. 围封与放牧管理对高寒草甸植物功能性状和功能多样性的影响[J]. 西北植物学报,2017,37(6):1216-1225.
[13]Bai Y F,Wu J G,Clark C M,et al. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:evidence from inner Mongolia Grasslands[J]. Global Change Biology,2010,16(1):358-372.
[14]Evans J R. Photosynthesis and nitrogen relationships in leaves of C3 plants[J]. Oecologia,1989,78(1):9-19.
[15]Garnier E,Cortez J,Billès G,et al. Plant functional markers capture ecosystem properties during secondary succession[J]. Ecology,2004,85(9):2630-2637.
[16]Mouchet M A,Villéger S,Mason N W H,et al. Functional diversity measures:an overview of their redundancy and their ability to discriminate community assembly rules[J]. Functional Ecology,2010,24(4):867-876.
[17]Westoby M,Falster D S,Moles A T,et al. Plant ecological strategies:some leading dimensions of variation between species[J]. Annual Review of Ecology and Systematics,2002,33(1):125-159.
[18]陈莹婷,许振柱. 植物叶经济谱的研究进展[J]. 植物生态学报,2014,38(10):1135-1153.
[19]Song C,Wang L,Tian H,et al. Effect of continued nitrogen enrichment on greenhouse gas emissions from a wetland ecosystem in the Sanjiang Plain,Northeast China:a 5 year nitrogen addition experiment[J]. Journal of Geophysical Research Biogeosciences,2013,118(2):741-751.
[20]李春丽,李 奇,赵 亮,等. 环青海湖地区天然草地和退耕恢复草地植物群落生物量对氮、磷添加的响应[J]. 植物生态学报,2016,40(10):1015-1027.
[21]李文娇,刘红梅,赵建宁,等. 氮素和水分添加对贝加尔针茅草原植物多样性及生物量的影响[J]. 生态学报,2015,35(19):6460-6469.
[22]Li W,Cheng J M,Yu K L,et al. Plant functional diversity can be independent of species diversity:observations based on the impact of 4-yrs of nitrogen and phosphorus additions in an alpine meadow,PloS One,2015,10(8):e0136040.
[23]Tilman D,Knops J,Wedin D,et al. The influence of functional diversity and composition on ecosystem processes[J]. Science,1997,277(5330):1300-1302.
[24]肖 迪,王晓洁,张 凯,等. 氮添加对山西太岳山天然油松林主要植物叶片性状的影响[J]. 植物生态学报,2016,40(7):686-701.
[25]万宏伟,杨 阳,白世勤,等. 羊草草原群落6种植物叶片功能特性对氮素添加的响应[J]. 植物生态学报,2008,32(3):611-621.
[26]杨 浩,罗亚晨. 糙隐子草功能性状对氮添加和干旱的响应[J]. 植物生态学报,2015,39(1):32-42.
[27]沈景林,孟 杨,胡文良,等. 高寒地区退化草地改良试验研究[J]. 草业科学,1999,16(3):4-7.
[28]Huberty L E,Gross K L,Miller C J. Effects of nitrogen addition on successional dynamics and species diversity in Michigan old-fields[J]. Journal of Ecology,1998,86(5):794-803.
[29]李文嬌,刘红梅,赵建宁,等. 氮素和水分添加对贝加尔针茅草原植物多样性及生物量的影响[J]. 生态学报,2015,35(19):6460-6469.
[30]Rajaniemi T K. Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses[J]. Journal of Ecology,2002,90(2):316-324.
[31]Li W,Wen S J,Hu WX,et al. Root-shoot competition interactions cause diversity loss after fertilization:a field experiment in an alpine meadow on the Tibetan Plateau[J]. Journal of Plant Ecology,2011,4(3):138-146.
[32]陈慧敏,石福习,杨桂生,等. 养分添加对三江平原沼泽化草甸植物群落组成和地上生物量的影响[J]. 生态学杂志,2016,35(6):1440-1446.
[33]Borer E T,Seabloom E W,Gruner D S,et al. Herbivores and nutrients control grassland plant diversity via light limitation[J]. Nature,2014,508(7497):517-520.
[34]Jutila H M,Grace J B. Effects of disturbance on germination and seedling establishment in a coastal prairie grassland:a test of the competitive release hypothesis[J]. Journal of Ecology,2002,90(2):291-302.
[35]Clark C M,Cleland E E,Collins S L,et al. Environmental and plant community determinants of species loss following nitrogen enrichment[J]. Ecology Letters,2007,10(7):596-607.
[36]Laughlin D C. Nitrification is linked to dominant leaf traits rather than functional diversity[J]. Journal of Ecology,2011,99(5):1091-1099.
[37]Macarthur R,Levins R. The limiting similarity,convergence,and divergence of coexisting species[J]. The American Naturalist,1967,101(921):377-385.
[38]Grime J P. Trait convergence and trait divergence in herbaceous plant communities:mechanisms and consequences[J]. Journal of Vegetation Science,2006,17(2):255-260.朱振華,张 艺,李小敏. 改性亚麻负载纳米铁对亚甲基蓝的吸附特性[J]. 江苏农业科学,2019,47(7):287-290.
