翘嘴鳜与斑鳜及正、反交F1代形态结构差异性
2019-09-17单建杰高四合杨广
单建杰 高四合 杨广
摘要:为比较翘嘴鳜和斑鳜自交后代及翘嘴鳜♀×斑鳜♂(正交F1代)、斑鳜♀×翘嘴鳜♂(反交F1代)杂交子一代形态结构特征差异,检测了相似生长环境条件的同批次4种鳜鱼群体的形态结构参数,利用单因子方差分析、聚类分析、判别分析等多元分析方法研究了4个群体间可数性状和可量性状的差异。结果表明,正反交F1代斑点特征介于亲本之间,更偏向于斑鳜,而与翘嘴鳜区别明显,其中正交F1代存在1条由背部向下延伸垂直于躯干的深褐色条带,而反交F1代无此性状。4个群体的背鳍(D)、胸鳍(P)、尾鳍(C)鳍条数目差异显著(P<0.05),存在重叠,不能作为区分依据,其中正交F1代的综合杂交指数(Hi1)更偏向母本,而反交F1代(Hi2)则居中间型。4个群体可量性状比值的单因子方差分析结果显示,除肝胰指数外,其他性状值间均存在显著差异(P<0.05),且正、反交F1代综合杂交系数(Hi1′、Hi2′)均偏向母本遗传。聚类分析结果显示,正、反交F1代形态最为接近,均偏向斑鳜,与翘嘴鳜差异较大。应用判别分析筛选出8个贡献较大的性状比值,构建判别式,判别准确率分别为 95.24%、81.58%、77.78%、84.72%,能够较好地判别4个群体。
关键词:斑鳜;翘嘴鳜;杂交;形态结构;可数性状;可量性状;判别准确率
中图分类号: S917.4 文献标志码: A 文章编号:1002-1302(2019)07-0193-04
翘嘴鳜(Siniperca chautsi)和斑鳜(S. scherzeri)同属于鲈形目鲈亚目鳜属,广泛分布于我国内陆水域,其肉质细嫩、味极鲜美,广受消费者的青睐,一直是我国重要优质名贵淡水鱼类。其中,翘嘴鳜是我国传统养殖品种,具有生长快、体型较大的优点,但抗病性较差,而且养殖期间病害频发[1]。与之相比,斑鳜的肉质更为细腻且抗逆性强、易驯食。此外,斑鳜的市场价格更高,出口需求大,但斑鳜生长速度较慢、养殖周期长,制约了其推广养殖[2-3]。
传统育种方式采用不同类型的亲本进行杂交,杂交后代会保留亲本的部分遗传性状[4-5],在杂交后代中筛选能兼顾两者优良性状的后代可以进一步促进鳜鱼养殖业的发展。本研究采用生长于我国北方地区的翘嘴鳜和斑鳜进行自交和杂交,培养子一代,比较子一代的形态结构特点,旨在为鳜鱼遗传育种研究提供参考。
1 材料與方法
1.1 试验鱼来源
本试验在天津市天祥水产养殖有限公司进行,试验用鱼为2016年经人工繁殖的子一代,养殖池塘条件、饵料种类及投喂比例基本一致,池塘为水泥池,水深2 m,面积0.13 hm2。养殖期间控制水温为20~23 ℃,溶氧量为7.26~8.87 mg/L,pH值为8.3~8.5。试验鱼规格见表1。
1.2 形态结构指标观察与测量
从4个群体中随机选取60~80尾健康鲜活鱼,观察其体型、体斑特征;计数背鳍(D)、胸鳍(P)、腹鳍(V)、臀鳍(A)、尾鳍(C)鳍条数等可数性状,测量全长、体长、头长、体高、头高、尾柄长、尾柄高、肠长等以及体质量、肝脏质量、内脏团质量等可量性状。
1.3 数据处理
为减少个体大小差异对形态结构研究的影响,将上述可量形态结构指标转化为形态比值作为分析对象,具体包括体长/全长、头长/体长、躯干长/体长、头高/体长、体高/体长、尾柄长/体长、尾柄高/体长、头高/体高、尾柄高/体高、肥满度、肠长指数、肝胰脏指数、内脏指数等指标。
1.3.1 方差分析 对翘嘴鳜、斑鳜及正反交F1代的5个可数性状以及13个可量性状比值进行Duncans多重比较,以检验不同群体形态是否有显著性差异。计算正、反交F1代形态指标的杂种指数Hi(hybrid index)[6]:
Hi=100×(Hi-Mi1)/(Mi2-Mi1);
式中:Hi表示杂种平均值;Mi1表示母本平均值;Mi2表示父本平均值。当4555为偏父本性状,Hi>100或Hi<0为超亲偏离性状。
1.3.2 聚类分析 利用SPSS 21.0对翘嘴鳜、斑鳜及正反交F1代13个性状比值进行系统聚类分析,获得欧式距离和聚类分析图。
1.3.3 判别分析 利用SPSS 21.0对翘嘴鳜、斑鳜及正反交F1代13个性状比值进行判别分析,根据Fisher线性判别式函数构建判别式,以函数最大值作为形态判别标准。
2 结果与分析
2.1 外形与体斑
翘嘴鳜口端位略上翘、梭形,背部隆起明显且宽厚,身体底色呈褐黄色和黑褐色,花斑呈上下不规则黑色条带状,奇鳍均呈黄色,其中尾鳍上有4~5列黑色斑点。斑鳜口端位、体修长、背部略隆起,体表底色呈褐黄色或棕色,背部及侧面有密集不规则圆斑,奇鳍上均分布2~3列黑色斑点。正、反交F1代体修长,背部平滑稍隆起,体侧底色为黄褐色和白色,体斑为黑色,体斑点数量与翘嘴鳜区别明显,偏向于斑鳜,但其形状和排布规则度均低于斑鳜,分布略稀疏;正、反交F1代尾鳍斑点排布同样偏向于斑鳜,为2~3列。此外,正交F1代体背部有1条垂直于躯干的条带,而反交F1代无此性状,见表2及图1。
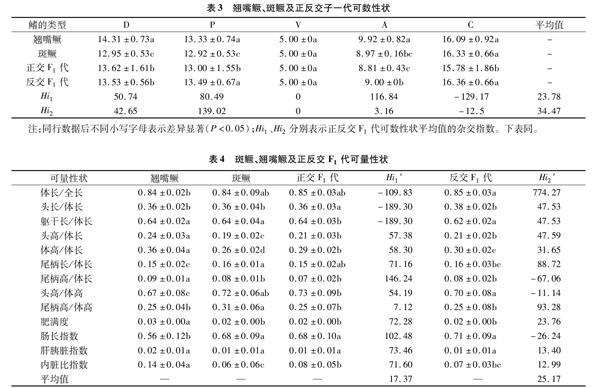
2.2 可数形状
本试验4种鳜鱼各鳍的鳍棘数相同,D、V、A鳍棘数分别为Ⅶ、Ⅰ、Ⅲ,P、C无硬棘。翘嘴鳜、斑鳜及正反交F1代各鳍软鳍条数见表3。正交F1代与亲本在D、P、C各鳍的鳍条数存在显著差异(P<0.05),其D、P鳍条数介于翘嘴鳜、斑鳜之间,C鳍条数少于亲本自交F1代,且D为中间性状(Hi1=50.74),P、A偏父本遗传(Hi1依次为80.49、116.84),C则超母本偏离(Hi1=-129.17)。
反交F1代与翘嘴鳜D、A鳍条数存在显著性差异(P<0.05),P、D、C鳍条数则与斑鳜差异显著(P<0.05);D、A、C鳍条数偏向母本斑鳜遗传(Hi2为42.65、3.16、-12.50),P鳍条数超父本翘嘴鳜偏离(Hi2=139.02),4种鱼的V鳍条数相同(Hi1、Hi2为0)。
可数性状综合平均杂交指数(Hi)显示,正反交F1代总体均偏向于母本遗传(Hi1=23.78,Hi2=34.47)。
2.3 可量性状
2.3.1 单因子方差分析 翘嘴鳜、斑鳜及正反交F1代可量性状的13个性状比值均值及差异显著性见表4。
正交F1代与翘嘴鳜、斑鳜相比,在头长/体长、躯干长/体长、头高/体长、体高/体长、内脏指数上均存在显著差异(P<0.05),其中头高/体长、体高/体长、尾柄长/体长、肥满度、肝胰脏指数、内脏指数属于偏父本性状(Hi1′分别为57.38、58.30、71.16、72.28、73.46、71.60),尾柄高/体长、肠长指数超父本偏离(Hi1′分别为146.24、102.48); 尾柄高/体高为偏母本性状(Hi1′=7.12),体长/全长、头长/体长、躯干长/体长均超母本偏离(Hi1′分别为-109.83、-189.30、-189.30);正交F1代共有8个性状偏向父本遗传,4个性状偏母本遗传。总体平均杂交指数(Hi1′)为17.37,偏向于母本翘嘴鳜遗传。
反交F1代与翘嘴鳜、斑鳜在头高/体长、体高/体长等比例性状均差异显著(P<0.05),其中体高/体长、肥满度、肝胰脏指数、内脏指数为偏母本性状(Hi2′分别为31.65、23.76、13.40、12.99),尾柄高/体长、头高/体高、肠长指数超母本偏离(Hi2′分别为-67.06、-11.14、-26.24);尾柄长/体长、尾柄高/体高偏向父本翘嘴鳜(Hi2′分别88.72、93.28),且头长/体长、躯干长/体长、头高/体长属中间性状(Hi2′分别47.53、47.53、47.59);反交F1代共有5个性状偏向父本遗传,7个性状偏母本遗传,在去除偏离较远的体长/全长后反交F1代平均杂交指数(Hi2′)为25.17,偏向于母本斑鳜。
2.3.2 聚类分析 翘嘴鳜、斑鳜与正反交F1代可量形状的聚类分析(图2)和欧式距离(表5)结果显示,正交F1代与反交F1代形态最为接近,先聚为一支,二者欧式距离最小,为 0.056;然后正反交F1代与斑鳜聚为一支,正交F1代与翘嘴鳜、斑鳜的欧式距离分别为0.157、0.085,略亲近于父本斑鳜;反交F1代与翘嘴鳜、斑鳜的欧氏距离分别为0.194、0.079,更亲近于母本斑鳜;且反交F1代与斑鳜距离近于正交F1代,说明反交F1代更偏向于斑鳜的形态;最后斑鳜与翘嘴鳜聚为一支,欧式距离为0.199。
2.3.3 判别分析 对所有形态比例参数进行判别分析,根据Fisher的线性判别式函数分类系数的大小, 筛选出贡献较大的8个性状比值,即体长/全长、头高/体长、体高/体长、尾柄高/体长、尾柄高/体高、肥满度、肠長指数、内脏指数,以x1~x8表示建立翘嘴鳜、斑鳜及正反交F1代的判别公式。
翘嘴鳜:y=1 780.637x1+798.633x2+1 731.380x3-4 741.449x4+1 641.705x5+3 893.264x6-32.305x7-141.254x8-1 184.233;
斑鳜:y=1 745.205x1+737.944x2+1 652.066x3-4 987.795x4+1 697.462x5+3 581.560x6+8.048x7-229.289x8-1 119.602;
正交F1代:y=1 745.029x1+765.001x2+1 679.437x3-4 861.206x4+1 650.884x5+3 350.194x6+2.689x7-216.1x8-1 122.23;
反交F1代:y=1 764.428x1+787.046x2+1 638.530x3-4 817.540x4+1 632.419x5+3 477.687x6+9.502x7-234.03x8-1 135.7。
将4个群体性状比值分别代入翘嘴鳜、斑鳜与正反交F1代4种鳜鱼判别函数中,以所得函数值最大作为判别标准,判别准确率分别为95.24%、81.58%、77.78%、84.72%,综合判别准确率为84.83%,能够较好地判别这4种群体。
3 讨论与结论
3.1 体斑、可数性状和可数性状偏向性分析
在杂交育种中,杂交后代的性状通常表现为不同性状的两亲本的中间态[7-8],本研究中正、反交F1代斑纹规则程度介于翘嘴鳜与斑鳜之间,与翘嘴鳜差异明显,更偏向于斑鳜,正交F1与反交F1斑纹的区别在于正交F1偏向母本翘嘴鳜鱼,具有1条从背部垂直躯干的长条斑纹;且正交F1存在2个可数性状偏向父本,1个性状超母本偏离,而反交F1代在可数性状上存在3个性状偏向母本遗传,1个性状偏父本遗传;结合可数性状平均杂交指数,可数性状综合平均杂交指数(Hi)显示,正交F1代、反交F1代总体均偏向于母本遗传(Hi1=23.78,Hi2=34.47)。翘嘴鳜、斑鳜及正反交F1代胸鳍鳍条数存在显著差异,然而具有重叠部分,不能作为区分4个群体的标准。
杂交子代在遗传性状等不同层面受双亲影响,且形态受母本遗传较多[9-11]。本研究中可量性状比值杂交指数(Hi)结果表明,正交F1代有8个偏父本斑鳜遗传,4个偏母本遗传,总体平均杂交指数(Hi1′)为17.37,总体偏向于母本翘嘴鳜遗传;反交F1代有7个偏向于母本遗传,2个偏向于父本遗传,3个偏向与中间性状,总体平均杂交指数(Hi2′)为 25.17,偏向于母本斑鳜遗传。由此看出,正交F1代、反交F1代分别遗传了双亲的性状,且综合杂交指数偏向母本遗传。
3.2 可量性状多元比较分析
外部形态特征是鱼类分类的依据[12],动物形态差异比较常采用多元分析法[13-14],本研究采用单因子方差分析、聚类分析和判别分析3种分析方法。3种分析方法从不同角度阐述了4个群体形态差异及遗传偏向性,综合单因子方差分析结果显示,正、反交F1代与翘嘴鳜、斑鳜在除肝胰脏指数之外的其他12个指标上均存在显著差异,差异主要体现在体长/全长、头高/体长、体高/体长、尾柄高/体长、尾柄高/体高、肥满度、肠长指数、内脏指数上,与判别分析中选用指标重叠,且判别分析进一步交代差异部位,为日后生产管理过程中品种判别提供理论基础;杂交指数交代了不同性状的亲本遗传偏向,可量性状比值综合杂交指数显示正交F1代、反交F1代均偏向母本遗传,与大多数鱼类形态研究结果[13-14]一致,然而聚类分析图及欧式距离显示,正、反交F1代与斑鳜距离更近,与翘嘴鳜距离略远,二者结论在正交F1上出现相悖,考虑原因为正交F1代中性状比值杂交指数偏向父本斑鳜个数(8个)较多,然而体长/全长、头长/体长、躯干长/体长超母本偏离(HI1′分别为-109.83、-189.30、-189.30),且有2个性状比值偏离较远,导致总体平均值偏向母本翘嘴鳜,2种分析从不同方面统计,得出了不同结果。正、反交F1代可量性状比值杂交指数均偏向于斑鳜的个数较多,性状个数与聚类分析结果一致。
综上所述,多元分析从不同角度对4个群体形态结构参数进行分析,且通过多元分析方法较好地展现出正、反交F1代与翘嘴鳜、斑鳜的差异性状及遗传的偏向性。正、反交F1代在不同层面遗传了亲本,部分性状超亲本偏离,单因子方差分析及判别分析展现了主要性状差异,且判别分析式为今后生产过程中品种判别提供理论基础,聚类分析显示正、反交F1代与斑鳜形态更为相似,杂交指数显示正、反交F1代存在较多个数的性状比值偏向于斑鳜。
参考文献:
[1]黄志坚,何建国. 鳜鱼疾病的研究概况[J]. 水产科技情报,1999(6):268-271.
[2]史建华,施顺昌,刘 峰,等. 杂交鳜与翘嘴鳜养殖比较研究[J]. 科学养鱼,2012(11):40-41.
[3]郑玉珍,王玉新,田功太,等. 斑鳜的生物学特性及繁殖技术[J]. 齐鲁渔业,2008(5):13-14.
[4]郭金涛,赵金良,甘远迪,等. 尼罗罗非鱼(♀)×萨罗罗非鱼(♂)杂交后代F1、F2形态性状的遗传与变异[J]. 中国水产科学,2014,21(2):275-282.
[5]葛玲瑞. 团头鲂(♀)×翘嘴红鲌(♂)杂交F1的生物学特征及遗传分析[D]. 长沙:湖南师范大学,2011.
[6]Matondo B N,Ovidio M,Poncin P,et al. Morphological recognition of artificial F1 hybrids between three common European cyprinid species:Rutilus rutilus,Blicca bjoerkna and Abramis brama[J]. Acta Zoologica Sinica,2008,54(1):144-156.
[7]吳水清,郑乐云,罗辉玉,等. 杂交石斑鱼(斜带石斑鱼♀×赤点石斑鱼♂)与其亲本形态性状比较研究[J]. 南方水产科学,2017(5):47-54.
[8]王 燕,张 勇,张海发,等. 两种杂交石斑鱼及其亲本的形态差异分析[J]. 水产学报,2014,38(6):778-785.
[9]孙翰昌,代 丽. 鲫(♀)×禾花鱼(♂)杂交及杂交F1、亲本的形态特征研究[J]. 淡水渔业,2011,41(1):48-52.
[10]李 迪. 草鱼(♀)×赤眼鳟(♂)F1及其亲本的肌肉营养和质构特性比较研究[D]. 长沙:湖南农业大学,2016.
[11]曹栋正,陈四清,严俊丽,等. 星突江鲽和石鲽正反杂交种的形态变异分析[J]. 中国水产科学,2016,23(4):871-881.
[12]孟庆闻. 鱼类分类学[M]. 北京:中国农业出版社,1996.
[13]唐瞻杨,林 勇,李莉萍,等. 四个尼罗罗非鱼引进种群的形态差异分析[J]. 西南农业学报,2012,25(2):718-722.
[14]霍堂斌,袁美云,马 波,等. 白斑狗鱼与黑斑狗鱼的形态差异与判别分析[J]. 大连海洋大学学报,2011,26(3):253-259.郑坚强,叶 豪,司俊玲,等. 响应面优化萃取宁夏枸杞类胡萝卜素工艺研究[J]. 江苏农业科学,2019,47(7):197-201.
