甘蓝型油菜主花序有效角果数QTL定位和候选基因筛选
2019-09-17耿鑫鑫曾焕黄伊雪
耿鑫鑫 曾焕 黄伊雪
摘要:以DH-7-9 (角果数多)× DH-G-42(角果数少)杂交后代连续自交的重组自交系(RIL)的190个家系为材料,在西宁和武汉2个环境下进行主花序有效角果数性状分析。结果表明,该RIL群体的主花序有效角果数表现出连续变异并且符合正态分布。利用已构建的遗传连锁图,结合2个年份2个环境下主花序有效角果数性状表型数据,采用WinQTL Cartographer 2.5软件的复合区间作图法(composite interval mapping,简称CIM)进行QTL 定位和效应估计。结果在2个环境下共检测到6个与主花序有效角果数、性状相关的QTL,单个QTL 可解释的表型变异范围为7.9%~24.39%。其中qESN-W-C5的贡献率达到24.39%,LOD值为12.97,位于C5染色体上,因此视其为该主花序有效角果数性状的主效QTLs。将拟南芥中已发表的角果数相关的3个基因与主效QTL置信区间对应的油菜基因组上104个基因进行同源比较分析,结果主效QTL区域内1个候选基因BnaC05g32840D与拟南芥角果数基因AT3G19820具有较高的同源性,推测其为油菜主花序有效角果数候选基因。
关键词:甘蓝型油菜;主花序有效角果数; QTL定位;候选基因
中图分类号: S634.303.2 文献标志码: A 文章編号:1002-1302(2019)07-0038-03
油菜是世界上分布最广、种植历史悠久的重要油料作物之一。甘蓝型油菜品质优、产量高,是我国油菜生产上的主要栽培类型[1]。菜籽油现在是国产食用植物油的第一大来源,在我国食用油市场中具有举足轻重的地位。目前,国内油料作物仅提供植物油消费总量的40%左右,约60%需进口,可见我国对国外的依存度越来越高,自给总量上严重不足[2]。双低菜籽饼粕每年为畜牧业提供超过600万t的饲料蛋白[3]。加拿大、澳大利亚、韩国、德国、法国、瑞典、奥地利等国确立为优势替代生物质能源(生物柴油) 原料作物,其中欧盟目前生产的生物柴油80%来自油菜原料(生产的油菜籽60%用于生产生物柴油)[4]。因此,提高油菜的产量成为了现在油菜育种的重要目标。
高产是作物育种的主要目标,产量性状是由微效多基因控制的数量性状,表现为连续变异,受环境的影响较大[5-6]。常规密度种植的油菜产量是由单株有效角果数、每角粒数以及千粒质量所构成,而这3个因素间既相互协同又相互制约。单株有效角果数是所有主要农作物产量构成的要素之一,其性状相对稳定。近年来随着农业机械化的发展,已经打破了常规密度人工种植油菜的传统观念,渐渐转向了机械化密植油菜的生产过程。机械化种植的油菜往往是密度高、分枝少,所以密植油菜主要是通过提高群体有效角果数来实现增产[7]。已有研究表明,适当增加密度可使主序在产量中的比例增加,在75万株/hm2时主序产量占单株产量的7305%[8]。而且油菜主序的2个主要性状主花序长度和主花序有效角果数一直以来都是影响单株产量的主要因素[9]。
综上所述,单株主花序有效角果数是影响油菜产量的重要因素。然而,目前为止,关于单株角果数数量性状基因座(quantitative trait locus,简称QTL)等方面的研究却严重不足。张书芬等在双低油菜细胞质雄性不育保持系1141B和双高恢复系垦C1构建的F2作图群体中检测出的3个单株角果数QTLs nsp4、nsp12和nsp2分别位于第4、第12和第2 连锁群上,分别解释变异方差的28.22%、11.97% 和7.73%[5]。易斌等利用中油821和保604为亲本材料,在相应DH系群体中将1个单株角果数QTL定位在第4连锁群上,可解释 17.42% 的表型变异[10]。孙美玉等以ZY036 × 51070 杂交构建的双单倍体(DH)为材料,在武汉、阳逻、青海和襄阳3年4点5个试验点共检测到10个与主花序有效角果数相关的QTLs,表型变异是9.33%~31.60%,在染色体A1、A5、C1和C9上的4个QTLs 都可以在2个不同的试验中重复检测到[11]。目前有关油菜单株角果数的研究基本停留在QTL定位分析水平,由于甘蓝型油菜基因组(AACC,异源四倍体)的复杂性,单株角果数相关基因的克隆与功能研究成果数量甚少。
本研究以甘蓝型油菜重组自交系(recombinant inbred lines,简称RIL)为材料,对油菜主花序有效角果数性状进行QTL定位。根据QTL置信区间对应的甘蓝型油菜基因组序列,找出该区间的全部基因,与拟南芥角果数相关基因进行同源比对并筛选候选基因。随着白菜、甘蓝及甘蓝型油菜全基因组序列陆续组装完成[12],后续精细定位和图位克隆相关基因及有关产量调控的分子机制研究必将达到一个全新的水平。
1 材料与方法
1.1 试验材料
以甘蓝型油菜品种DH-7-9(主花序有效角果数多年多点至少80个)和DH-G-42(主花序有效角果数为多年多点至多58个)为材料进行杂交得到F1植株,构建了包含190个株系的重组自交系群体。本试验利用该群体进行田间试验、主花序有效角果数性状统计和QTL分析。
1.2 田间试验与主花序有效角果数性状考察
将重组自交系群体于2013年9月种植于中国农业科学院油料作物研究所武汉市武昌区试验田,2013年5月及2014年5月种植于青海省西宁市青海大学油菜繁育中心试验田。所有田间试验均按3个重复分区种植,每小区3 行,行距 40 cm,株距20 cm。按常规生产方式进行田间管理。2014年4月(武汉)、2013年10月(西宁)和2014年10月(西宁)收获时期从每个小区内随机选取5~10 株,用于主花序有效角果数性状的测定(每1株油菜植株主花序上能够结1粒及以上饱满种子的角果数)。
1.3 主花序有效角果数的QTL定位
在利用F2代群体已构建好的遗传连锁图谱[6]的基础上,进行主花序有效角果数QTL定位。该连锁图包含228个标记,20个连锁群,图谱总长为1 546.6 cM。用WinQTL Cartographer 2.5软件,采用复合区间作图(CIM)的方法[13-14]定位该重组自交系群体的主花序有效角果数QTL。选取 1 cM 的步长,在α=0.05的水平上,利用Permutation检验法重复检验1 000次。将LOD阈值设定为2.5来确定QTL在染色体上的位置及数目。QTL命名参照McCouch等文献报道的相关规则[15],用SPSS 16.0软件(SPSS,Chicago,IL,USA)统计分析表型数据。
1.4 候选基因筛选
通过对TAIR网站(http://www.arabidopsis.org/)以及已发表的文章[16]中拟南芥相关基因功能信息进行分析共获得了3个角果数相关基因。在NCBI上将此3个拟南芥角果数相关基因序列进行下载,然后与检测到的主效QTL置信区间对应的甘蓝型油菜基因组序列[12]进行BlastN比对,并进行候选基因的筛选。
2 结果与分析
2.1 双亲和RIL群体主花序有效角果数的表型统计及与其他农艺性状的相关性
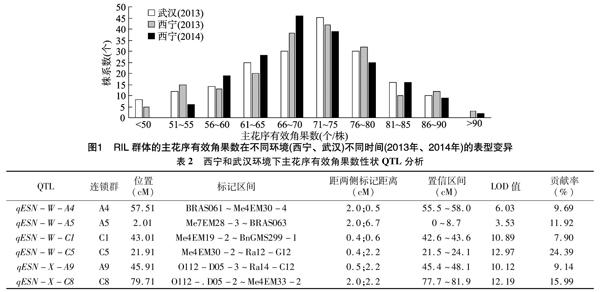
对双亲和RIL群体主花序有效角果数表型进行統计,结果表明,DH-G-42-8和DH-7-9-6主花序有效角果数差异很大,并且两亲本间具有极显著性差异(表1)。统计分析表明,主花序有效角果数在重组自交系群体中呈现超双亲分离和连续性分布并且符合正态分布(图1),说明主花序有效角果数是由多个基因控制的数量性状,适合于QTL 分析。
2.2 主花序有效角果数性状QTL定位
通过QTL Cartographer 2.5复合区间作图法进行QTL分析,在武汉和西宁2个环境下共检测到6个QTLs与油菜主花序有效角果数性状相关,分别位于A4、A5、A9、C1、C5以及C8染色体上(表2),分别命名为qESN-W-A4、qESN-W-A5、qESN-W-C1、qESN-W-C5、qESN-X-A9和qESN-X-C8。3个QTLs(分别为qESN-W-A5、qESN-W-C5和qESN-X-C8)的贡献率都大于10%,其中qESN-W-C5的贡献率达到24.39%,LOD值为12.97,因此视其为该主花序有效角果数性状的主效QTL(图2)。
2.3 候选基因筛选
通过生物信息学分析,将该主效QTL置信区间(约0.94 Mb)与甘蓝型油菜基因组序列比对, 得到共104个注释基因。将这些基因与已知的拟南芥3个角果数相关基因(AT1G68725、AT2G33150和AT3G19820)进行同源比对,结果发现主效QTL置信区间内有1个基因(BnaC05g32840D)与拟南芥角果数基因具有很高的同源性(88.8%)。拟南芥同源角果数基因为AT3G19820,编码DWF1/DIM,在油菜素类固醇(BR)的生物合成中催化早期BR前体24-亚基的合成。油菜素类固醇会影响细胞的伸长,其突变体具有矮化表型,而 DWF1是1个Ca2+依赖性钙调素结合蛋白。
3 讨论与结论
本研究所选用的2个亲本材料主花序有效角果数表型存在极显著差异,RIL群体表型呈现连续分布,所选用的甘蓝型油菜遗传图谱总长1 546.6 cM,标记间的平均图距为 6.78 cM,图谱质量可靠。因此,这2个亲本材料及图谱适合研究主花序有效角果数相关遗传分析。
在国内外的报道中,孙美玉等以ZY036×51070杂交构建的DH群体为材料,3年4点5个试验共检测到10个与主花序有效角果数相关的QTLs,分别位于A1、A3、A5、C1、C4、C6、C9,解释性状表型变异是933%~31.60%[11]。高必军利用中双4号×H228构建RIL群体,共检测到主花序有效角果数5个QTLs,分别位于A6、C2、C3连锁群,解释性状变型变异是11.2%~25%[17]。吴建忠检测到3个与主花序有效角果数相关的QTLs,分别位于A8和C4上,解释性状表型变异507%~9.45%[18]。本研究一共检测到6个主花序有效角果数QTLs,分别位于A4、A5、A9、C1、C5和C8上,贡献率在7.9%~24.39%之间,LOD值为3.53~12.97。跟前人的研究结果相比,本研究在A4、A9、C5和C8新发现了主花序有效角果数QTLs。一般认为,贡献率在10%以上的QTL为主效QTL,LOD值越大,QTL的准确率越高。其中qESN-W-C5的贡献率达到24.39%,LOD值为12.97,为所定位的6个QTLs中的最大值,因此视其为该主花序有效角果数性状的主效QTL。
QTL与环境之间存在密切的关系。本研究中共检测到的6个QTLs是在2个年份2个环境中检测到的,并没有可以在2年相同环境下同时检测到,产生这些结果的原因:一是油菜主花序有效角果数性状在不同环境条件下的不稳定表达;二是不能重复检测到有可能代表了该环境条件下的特异表达基因;三是群体中的个体可能受到环境条件的影响程度不一造成的误差。
通过对QTL置信区间与基因组序列比对进行候选基因预测,可以比较简便快速地验证QTL定位的准确性。在C5染色体上的候选区域与报道的拟南芥角果数相关基因进行比对,发现了1个同源性较高的候选基因BnaC05g32840D,其编码DWF1/DIM,在油菜素类固醇(BR)的生物合成中催化早期BR前体24-亚基的合成。油菜素类固醇会影响细胞的伸长,其突变体具有矮化表型,从而调控植株的有效角果数数量。后续将对该候选基因进行相关功能验证,并同时利用主花序有效角果数主效QTL附近的分子标记逐步构建目的基因的近等基因系,通过精细定位和图位克隆最终获得控制该性状的主效基因。
参考文献:
[1]张亚宏,雷建明,张 岩,等. 浅析强冬性甘蓝型油菜的生产与发展潜力[J]. 农业科技通讯,2010(1):103-106.
[2]王汉中. 我国油菜产业发展的历史回顾与展望[J]. 中国油料作物学报,2010,32(2):300-302.
[3]沈金雄,傅廷栋,涂金星,等. 中国油菜生产及遗传改良潜力与油菜生物柴油发展前景[J]. 华中农业大学学报,2007,26(6):894-899.
[4]徐桂转,张百良. 生物柴油的研究与进展[J]. 华中农业大学学报,2005,24(6):644-650.
[5]张书芬,傅廷栋,朱家成,等. 甘蓝型油菜产量及其构成因素的QTL定位与分析[J]. 作物学报,2006,32(8):1135-1142.
[6]朱恒星,闫晓红,方小平,等. 甘蓝型油菜千粒质量性状的QTL初步定位研究[J]. 植物遺传资源学报,2012,13(5):843-850.
[7]李孟良.不同密度对直播油菜生长及产量的影响[J]. 安徽科技学院学报,2011,25(1):23-26.
[8]张宇文,郭亚茹. 油菜主序优势及其利用初析[J]. 西北植物学报,1996,16(6):126-131.
[9]宋 稀,刘凤兰,郑普英,等. 高密度种植专用油菜重要农艺性状与产量的关系分析[J]. 中国农业科学,2010,43(9):1800-1806.
[10]易 斌,陈 伟,马朝芝,等. 甘蓝型油菜产量及相关性状的QTL分析[J]. 作物学报,2006,32(5):676-682.
[11]孙美玉,华 玮,刘 静,等. 甘蓝型油菜主花序有效角果数QTL定位[J]. 中国油料作物学报,2013,35(1):1-7.
[12]Chalhoub B,Denoeud F,Liu S Y,et al. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome[J]. Science, 2014,345(6199):950-953.
[13]Zeng Z B. Theoretical basis for separation of multiple linked gene effects in mapping quantitative trait loci[J]. Proceedings of the National Academy of Sciences of the United States of America,1993,90(23):10972-10976.
[14]Zeng Z B. Precision mapping of quantitative trait loci[J]. Genetics,1994,136(4):1457-1468.
[15]McCouch S R,Cho Y G,Yano M,et al. Report on QTL nomenclature[J]. Rice Genet Newsl,1997,14(11):11-13.
[16]Shi J,Li R,Qiu D,et al. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus[J]. Genetics,2009,182(3):851-861.
[17]高必军. 甘蓝型油菜napin基因启动子的克隆与几个重要农艺性状的初步QTL定位[D]. 雅安:四川农业大学,2007:1-121.
[18]吴建忠. 甘蓝型油菜结实相关性状分析及QTL定位[D]. 武汉:华中农业大学,2010:1-60.张 宁,尹美强,谭青青,等. 苦参转录组SSR位点及基因功能注释分析[J]. 江苏农业科学,2019,47(7):41-44.
