北方寒区日光温室冬季基质袋培番茄蒸腾量模拟
2019-09-16罗新兰佟国红张函奇李英歌尹佳琪杨丽桃
罗新兰,王 淼,4,佟国红,张函奇,李英歌,尹佳琪,杨丽桃
(1.沈阳农业大学农学院,辽宁 沈阳 110866; 2.沈阳农业大学水利学院,辽宁 沈阳 110866;3.内蒙古自治区气候中心,内蒙古 呼和浩特 010051;4.锦州市气象局,辽宁 锦州 121000)
冬季日光温室是个相对封闭的生产环境,室外温度很低,室内很少通风,作物蒸腾是作物冠层与室内空气之间进行水汽、能量交换的主要途径,会直接影响室内温湿度和能量平衡,作物蒸腾消耗热能降低室内空气温度,释放水汽增加室内湿度,室内湿度的改变又会影响作物蒸腾及病虫害的发生。人工测量作物蒸腾费时费力,利用彭曼公式计算作物需水量是联合国世界粮农组织推荐的一种计算方法[1],Monteith在1963年对Penman方程进行改进,提出Penman-Monteith(P-M)方程,其以能量平衡为依据,加之水汽扩散为基础,适用于任何作物蒸腾量的直接计算[2-3]。Moritlle等[4]验证了温室条件下采用P-M方程计算作物蒸腾量的可行性。
Boulard等[5]及Demrati等[6]提出基于P-M方程,综合了室内能量平衡和室外气象数据建立了温室作物蒸腾模型,结果显示当温室处于通风状态时,蒸腾模型模拟结果良好。罗卫红等[7],汪小旵等[8]应用P-M方程分别模拟南方冬季和夏季现代化温室黄瓜蒸腾情况,结合室内净辐射及饱和水汽压差(VPD)分析蒸腾速率的日变化,在冬季,作物蒸腾速率日变化主要取决于太阳辐射的日变化,而与VPD的日变化关系不大;在夏季,作物蒸腾速率对太阳辐射和VPD的变化敏感性相同。番茄是温室内的主要作物,专家学者为探寻温室条件下番茄的蒸腾规律及影响因素,在塑料大棚[5]、玻璃温室[9]、防虫遮阳网温室[10]、日光温室[11-16]环境下,开展了深入研究。由于影响蒸腾的因素较多且很多参数很难精确测定,目前还缺乏通用的、精准的、用于温室不同条件下的番茄蒸腾模型。
温室中基质袋培可避免土壤的连作障碍和土传性病虫害的蔓延,具有管理方便、原料资源丰富、经济环保等特点。北方寒区日光温室冬季生产基本无通风,室内弱光、高湿、低温及低风速,为确定该环境下单株番茄蒸腾量模拟模型,本研究以P-M方程为基础,采用室内单株尺度下气象环境特征对应的边界层空气动力学阻力、气孔平均阻力反演模型并引入土壤热通量参数,得到一个基于温室内气象数据(温度、相对湿度、太阳辐射等)和植株生长指标(茎粗、单株叶面积指数等)计算地面覆盖塑料薄膜的北方寒区日光温室冬季基质袋培单株番茄蒸腾量的方程。根据伍德林等[17]研究显示作物吸收水分的近99.8%用于植株蒸腾作用,因此可用插针式茎流计测定植株茎流量作为实测值对单株植株蒸腾模拟结果进行验证。单株番茄蒸腾量的模拟可为北方寒区冬季番茄滴灌的灌溉量确定提供参考。
1 材料与方法
1.1 试验概况
试验于2017-12-11—2018-01-03在沈阳农业大学北山科研基地日光温室(123°5′E,41°8′N)内进行。温室方位南偏西7°,东西长度60 m,跨度7.5 m,脊高3.5 m,北墙高2.3 m,墙体厚45 cm,北墙外侧设有冷棚,试验期间揭帘时间8∶30,盖帘时间14∶30。
供试番茄品种为适于北方日光温室长季节栽培的圣罗兰。植株采用南北成行基质袋培,每行铺有一条滴灌管,各滴头与各植株相对应,滴头出水口插于栽培袋中间位置(晴天时于9∶00、12∶00、14∶00分别灌溉100 ml·株-1·次-1),基质水分控制在田间持水量80%以上,地面覆盖塑料薄膜(无裸露土壤),行距1 m,株距20 cm,共定植920株,定植密度为3.38株·m-2。选择长势良好的3株番茄作为测试样本。温室顶部通风口可人工开启,晴天中午室内温度超过30℃时通风口打开10 cm进行通风,平均通风时间为15 min。其余时间处于密闭状态,试验期间植株处于成熟采摘期,茎粗1.41~1.48 cm、株高188.3~193.6 cm。
1.2 观测方案及布置
室内中部安装HOBO自动气象站以获取试验所需基本气象数据(辐射、温度、相对湿度、土壤含水量等);番茄群体中间安装北京雨根公司的环境及茎流监测仪,测定项目包括:植株上方的太阳净辐射、植株的茎流值、空气温湿度、叶温(用与叶片相同颜色的绿色别针将热电偶与叶片固定);采用热球风速仪测量植株上方微风速。试验期间24 h连续测试,取样间隔为10 min,蒸腾速率估算中用到的相关气象要素观测项目及仪器位置见表1。
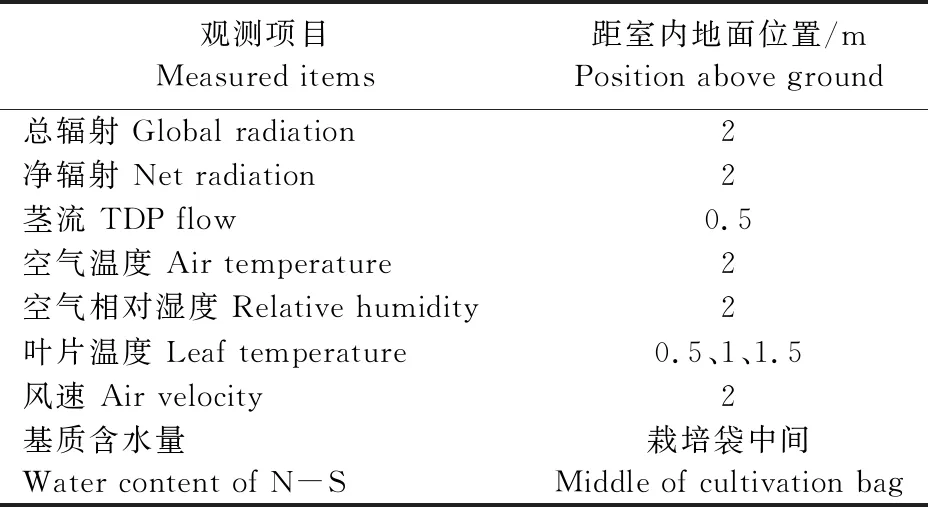
表1 日光温室内气象要素观测项目及仪器位置
本试验采用插针式茎流计测量番茄蒸腾速率作为实测值。用公式(1)对每株实测数据ΔT做统计学分析,修正不合理的ΔT值
(1)
式中,ΔT′为两探针之间温差修正值;ΔT为实测两探针之间温差;ΔT5为与ΔT相对应滑动长度为5的ΔT原序列的滑动平均值,SE为相应滑动标准差。
将修正后的数据采用以下计算式得出茎流速率。
根据Granier提出的基本计算方法[18],其计算植株茎流速率的公式如下:
Fd=118.99×10-6K1.231
(2)
式中,Fd为茎秆液流速率(m·s-1);K为系数。
(3)

将上述液流速率转换为质量形式,即[19]:
Fs=3600×FdρAs
(4)
式中,Fs为质量形式茎秆液流速率(g·h-1);Fd为茎秆液流速率(m·s-1);ρ为水密度(g·m-3);As为茎秆横截面积(m2)。
将Fs带入下式可计算出单株植株实测蒸腾速率值[20-21]:
(5)
式中,Tf为单株植株蒸腾速率(mm·h-1);La为植株监测探针上方的叶面积(m2);LAI为单株叶面积指数(某株植株叶面积/某株植株占地面积)。
1.3 分析方法
1.3.1 单株叶面积指数(LAI) 叶面积采用主干上主叶长宽系数法计算,测量主叶的叶长(包括叶柄长度)和最大叶宽,计算公式如下[22]:
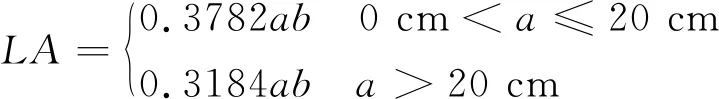
(6)
式中,LA为叶面积(cm2);a为叶片长度(cm);b为叶片宽度(cm)。LAI=LA/单株植株占地面积。
1.3.2 消光系数(K) 根据比尔-朗伯特(Beer-Lambert)定律得到:
(7)
式中,LAI为单株叶面积指数;I,I0分别为单株尺度的地面和植株上方的总辐射(W·m-2)。计算每日9∶00—14∶00内每10 min的消光系数,根据冯新灵等[23]研究建议剔除不合理的消光系数数据(即K>1的数据),再以日为单位将合理数据均值后作为每日的消光系数。
1.3.3 采用Penman-Monteith方程模拟日光温室基质袋培单株植株蒸腾量 在考虑土壤热通量、并结合密闭环境下的边界层空气动力学阻力计算式及气孔平均阻力反演式,Penman-Monteith方程计算温室内单株植株蒸腾量的基本式可转换为[24-26]:
(8)

R'n=Rn(1-e-K·LAI)
(9)
式中,Rn为单株植株上方净辐射(W·m-2);K为消光系数;LAI为单株叶面积指数。
根据Goudriaan和Van[27]:
(10)
式中,ta为室内空气温度(℃);Δ为饱和水汽压随温度变化曲线斜率(kPa·℃-1)。
(11)
联合国粮食及农业组织(简称FAO)推荐土壤热通量G白天采用0.5Rn,夜间采用0.1Rn[3],本文根据北方寒区日光温室揭、盖帘时间对应的温室内日照时数对G进行修正,白天采用0.35Rn,夜间采用0.13Rn,后文对此取值进行敏感性检验。
1.3.4 边界层空气动力学阻力(ra) 叶片与周围环境热交换形式与ra大小密切相关[28],风是作物与周围环境进行物质和能量交换的动力来源,其大小决定热交换速率,Stanghellini[29]研究表明在任何温室中均会发生以混合对流形式存在的对流热交换,即自由对流与强迫对流的混合,当室内风速低于0.2 m·s-1时,强迫对流对ra影响很小,可忽略。
根据Campbell[30]研究结果,北方寒区冬季日光温室密闭条件下由叶温与周围空气温度差值引起的自由对流对ra起决定作用,计算公式如下:
(12)
(13)
式中,ra1为单一叶片边界层空气动力学阻力(s·m-1);d为叶片特征长度(m)(本试验取值0.15 m);Tl为叶片温度(℃);Ta为周围空气温度(℃);ra为单株植株边界层空气动力学阻力(s·m-1);LAI为单株叶面积指数。
1.3.5 单株植株对水汽的阻力(气孔平均阻力,rc) 作物通过叶片气孔与周围环境进行显热交换和水分传输,交换和传输过程中受气孔阻力控制,由于影响温室内气孔平均阻力的主要因素包括室内总辐射、温度、饱和水汽压差等气象因子,因此可以通过反演的方式计算出植株样本a的rc值。rc计算表达式如下所示:
(14)
式中,λE为植株a的实测蒸腾潜热值(W·m-2);ra为植株a的边界层空气动力学阻力(s·m-1);其余物理量为植株a单株尺度对应的数值且意义同前。
2 结果与分析
将公式(14)计算的rc值、植株b、c相对应的ra值及生理指标(LAI)值代入公式(8),计算b、c株蒸腾速率模拟值,并与其相应的实测值比较分析,检验P-M方程模拟效果。由1.3.1计算的试验期间LAIa=1.18,LAIb=1.05,LAIc=1.08。
2.1 日光温室内单株植株边界层空气动力学阻力与气孔平均阻力的变化规律
选取试验期间有典型天气的2017年12月13—25日(其中13日与17日为雾霾天、14日与24日为阴天,其余时间为晴天)连续13 d的测试值计算植株a的边界层空气动力学阻力(ra)值、气孔平均阻力(rc)值,ra与rc在不同天气变化情况如图1所示。
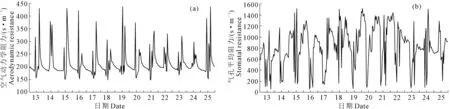
图1 冬季日光温室内单株番茄边界层空气动力学阻力(a)与气孔平均阻力(b)变化规律(2017年12月13—25日)Fig.1 Variation of aerodynamic resistance and leaf stomatal resistance of single tomato plant in CSG during winter (Dec.13-25, 2017)
由图1(a)可见,ra晴天白天的波峰在10∶00,变化范围为259~438 s·m-1;波谷在15∶00,变化范围为147~198 s·m-1。阴天白天波峰在11∶00,变化范围为297~365 s·m-1,波谷在13∶00,变化范围为211~243 s·m-1。这是由于10∶00时|Tl-Ta|最小,放帘后半小时左右|Tl-Ta|达最大值,在中午|Tl-Ta|≈5℃,ra≈180 s·m-1,阴天时太阳辐射弱,室内温度变化慢,在11∶00左右|Tl-Ta|达最小值,13∶00左右|Tl-Ta|即达最大值。
由图1(b)可见,rc晴天、阴天白天的波峰在7∶00,变化范围分别为820~1 506、749~1 151 s·m-1,波谷在11∶00—13∶00,变化范围分别为69~205、132~208 s·m-1。这是因为rc主要取决于室内总辐射的变化,总辐射越高,rc越小,冬季8∶30-14∶30为揭帘时间室内总辐射为正值,其余时间室内总辐射为0,日出后室内总辐射迅速上升,rc随即下降,11∶00-13∶00单株植株分得的总辐射在一天中最大值约为100 W·m-2,rc达到最低值,约为158 s·m-1。
2.2 日光温室内单株植株蒸腾速率模拟值与实测值比较
将室内单株尺度的气象要素及相关公式带入公式(8)计算植株b、c的模拟值并与其相对应的茎流计实测值进行逐时蒸腾速率对比,结果如图2所示。

图2 冬季日光温室内单株番茄逐时蒸腾速率模拟值与实测值对比(2017年12月13—25日)Fig.2 Comparison between simulated and measured hourly transpiration rate of single tomato plant in CSG during winter (Dec.13-25, 2017)
由图2可知,逐时蒸腾速率模拟值的最大值在晴天约为0.06 mm·h-1,阴天约为0.02 mm·h-1,晴天、阴天均与实测值相当,而雾霾天实测值超过模拟值约为0.015 mm·h-1,误差较大。这是因为在晴天、阴天室内各气象数据均是较规律的变化,而雾霾天随大气透明度的变化,室内气象数据发生不规律变化。由于北方冬季夜间或早晚室外温度低时温室覆保温被以保证室内温度满足植株生理活动的最低要求,早、晚覆保温被期间室内总辐射为0,蒸腾速率很小甚至为0,因此,为进一步说明模型拟合精度,计算整个试验期间所有晴天与阴天(除去所有雾霾天)植株b、c的模拟值与实测值24 h的累计蒸腾量,结果如图3所示

图3 冬季日光温室内单株番茄晴天与阴天24 h累计蒸腾量 模拟值与实测值对比Fig.3 Comparison between simulated and measured accumulated transpiration values of single tomato plant in CSG during winter
由图3可以看出植株b、c的24 h累计蒸腾量模拟值与实测值一致性较好,植株b的R2=0.9228、截距为0.0046 mm·d-1;植株c的R2=0.9594、截距为0.0115 mm·d-1。单株日累计蒸腾量晴天变化范围为0.22~0.36 mm·d-1,阴天为0.08~0.15 mm·d-1,用1∶1检验法对植株b、c累计蒸腾量模拟值与实测值对比分析结果表明:24 h日累计蒸腾量均匀分布于y=x直线两侧,为进一步说明误差大小,分别计算两株植株整个试验阶段24 h累计蒸腾量模拟值与实测值的平均相对误差(MRE)和平均绝对误差(MAE),计算表达式为:
(15)
(16)
式中,n为测试蒸腾量天数;yi为第i天蒸腾量模拟值;xi为第i天蒸腾量实测值。结果如表2所示。
由表2可知整个试验阶段植株b、c 24 h累计蒸腾量的MRE和MAE分别为9.46%、10.98%和0.02 mm·d-1、0.02 mm·d-1。晴天、阴天约占总试验天数的9/10,说明使用P-M方程模拟北方寒区冬季日光温室基质袋培单株番茄蒸腾量(即需水量)是可行的。
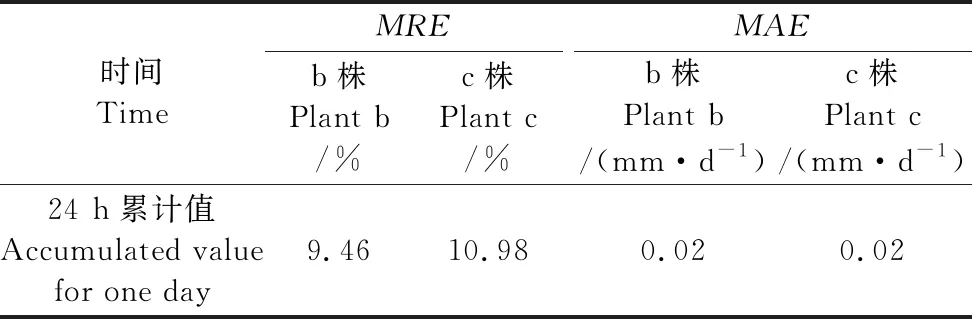
表2 冬季日光温室内番茄植株累计蒸腾量误差分析
2.3 日光温室内单株植株上方净辐射、空气饱和水汽压差、蒸腾速率的日变化规律
净辐射的大小直接关系到蒸腾作用的强度,温室内饱和水汽压差(以下简称VPD)直接影响植物气孔闭合程度,从而控制作物蒸腾过程和水分利用效率,两者作为温室作物蒸腾的主要影响因子,共同决定温室番茄植株的蒸腾速率。选取试验期间每天9∶00-15∶00植株b上方净辐射、距地面2 m处的参考高度VPD及其蒸腾速率的关系如图4所示。
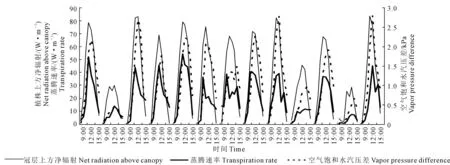
注:植株蒸腾速率(mm·h-1)转换为蒸腾作用带走的热量(W·m-2)。 Note: Plant transpiration rate (mm·h-1) was converted to heat carried by transpiration (W·m-2).图4 冬季日光温室内单株番茄上方净辐射、空气饱和水汽压差(VPD)与蒸腾速率的日变化规律Fig.4 Variation of net radiation, vapor pressure difference(VPD), and transpiration rate of single tomato plant in CSG during winter
由图4可看出,冬季日光温室单株番茄蒸腾速率与其上方净辐射保持很好的一致性,日最大值均出现在12∶00左右且几乎保持同步,而VPD日最大值出现时间滞后于前两者1 h左右。例如12月23日,净辐射突然下降时,蒸腾速率随即下降,而VPD在1 h后明显降低。由图1b可看出9∶00—15∶00气孔阻力维持在较低值,由此可得出在冬季,日光温室单株番茄蒸腾速率的大小主要取决于其上方净辐射的日变化规律和大小,而室内VPD的变化和大小对其起次要作用,这与罗卫红等[7]关于南方冬季现代化温室内净辐射、VPD与黄瓜蒸腾速率的日变化规律的研究趋势相同。
2.4 日光温室内单株植株上方净辐射与蒸腾速率的关系
净辐射是温室作物蒸腾的源动力,根据2.3中分析可知冬季日光温室单株番茄蒸腾速率变化规律与其上方净辐射变化规律一致,为探明室内植株上方净辐射对蒸腾速率的影响,根据试验数据得出植株b上方净辐射与蒸腾速率关系如图5所示。
由图5可知两者呈线性关系,约有43.5%的植株上方净辐射通过植株蒸腾作用转化为潜热,而根据罗卫红等[7]对南方现代化温室黄瓜冬季蒸腾的研究结果,平均有46%作物冠层上方净辐射通过作物蒸腾作用转化为潜热,本文试验结果比其低,这是因为测试期间晴天中午平均室内总辐射为165.6 W·m-2,室内平均温度为21.6℃(南方现代化温室黄瓜冬季蒸腾试验期间相应值为187 W·m-2,26.2℃),室内总辐射与温度均较低使室内VPD维持较低水平,冬季室内无通风,风速极低甚至为0,导致蒸腾作用较弱,且番茄叶片蒸腾强度低于黄瓜叶片,进而使植株上方净辐射通过蒸腾作用转化为潜热的比例明显降低。
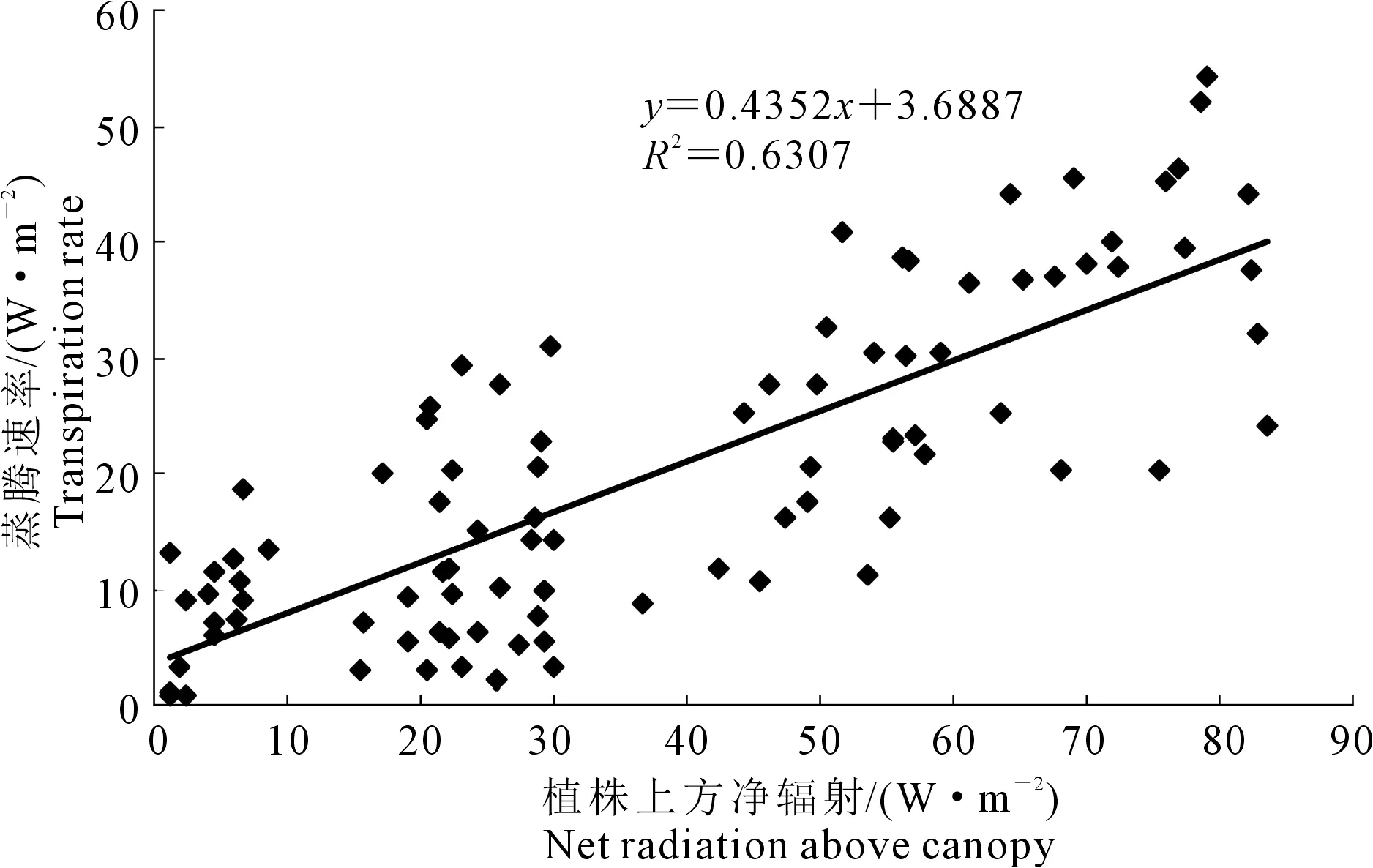
注:植株蒸腾速率(mm·h-1)转换为蒸腾作用带走的热量(W·m-2)。 Note: Plant transpiration rate (mm·h-1) was converted to heat carried by transpiration (W·m-2).图5 冬季日光温室内单株番茄上方净辐射与蒸腾速率的关系Fig.5 Relationship between the net radiation above plant and the measured transpiration rate ofsingle tomato plant in CSG during winter
2.5 日光温室内土壤热通量的敏感性检验
温室地面在白天、夜间与温室内部进行能量交换,为了检验土壤热通量的变化对蒸腾速率的影响,以12月20日9∶00—22∶00、12月21日0∶00—7∶00(本研究土壤热通量白天采用0.35Rn,夜间采用0.13Rn)的植株b逐时蒸腾速率模拟值为例,分别计算白天采用0.25~0.5Rn、步长为0.05Rn,夜间采用0.1~0.15Rn、步长为0.01Rn进行敏感性检验,计算结果如图6所示。
由图6分析可知,尽管18∶00植株b的模拟值对G变化敏感性较强,但由于此时蒸腾速率较低,因此变化值的大小对日累计蒸腾量的影响效果不大,白天G/Rn每增加0.05时,对单株植株蒸腾速率影响最大时间段为10∶00—11∶00,蒸腾速率平均减少了5.46%,影响最小时间段为14∶00—15∶00,蒸腾速率平均减少了1.55%;夜间G/Rn每增加0.01时,仅在18∶00时蒸腾速率增加了4.75%,其余时间均增加2.25%。土壤热通量变化对逐时蒸腾速率影响仅在白天一段时间略超过5%,其余时间均低于5%,由此可见在本试验条件下土壤热通量对逐时蒸腾速率的影响不明显。
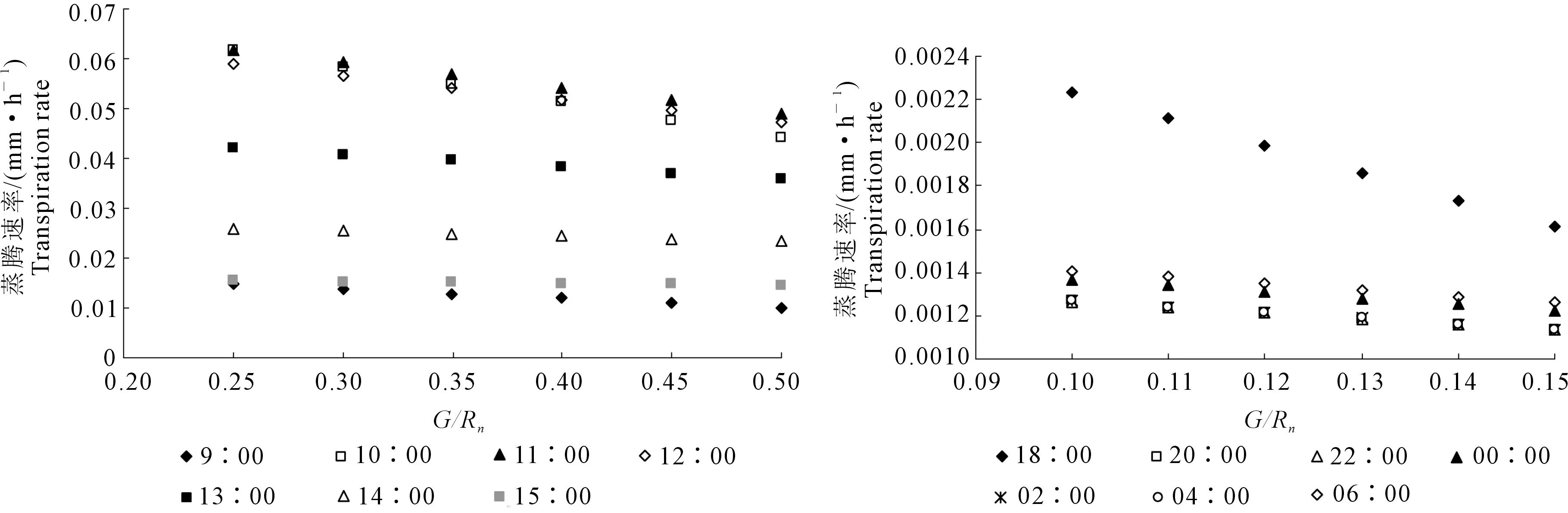
图6 冬季日光温室内土壤热通量敏感性分析Fig.6 Sensitivity analysis of soil heat flux in CSG during winter
3 结论与讨论
3.1 结论
(1)北方日光温室冬季密闭环境中晴天、阴天单株番茄边界层空气动力学阻力ra的平均变化范围分别为147~438、211~365 s·m-1;气孔阻力rc平均变化范围分别为69~1 506、132~1 151 s·m-1。
(2)北方日光温室冬季密闭环境中基于P-M方程模拟单株番茄逐时蒸腾速率是可行的:检验模型的b、c植株日累计模拟值与茎流计实测值间的R2>0.9,标准差SE≈0.01 mm·d-1,b、c株模拟值与实测值的平均相对误差分别为9.46%、10.98%;晴天、阴天中午的平均值分别为0.06、0.02 mm·h-1,日蒸腾总量平均值分别为0.29、0.12 mm·d-1。
(3)在北方冬季低温高湿的环境下,作物蒸腾速率主要取决于植株上方净辐射的变化,植株上方净辐射与蒸腾速率的最大值均出现在12∶00左右,距离地面2 m处参考高度VPD最大值出现时间约13∶00。植株上方净辐射量的43.5%可通过蒸腾作用转化为潜热。
(4)P-M方程估算单株植株逐时蒸腾速率时考虑了土壤热通量一项,研究发现晴天白天G/Rn每增加0.05对蒸腾速率影响最大时间段为10∶00—11∶00,蒸腾速率减少5.46%,影响最小时间段为14∶00—15∶00,蒸腾速率减少1.55%;夜间G/Rn每增加0.01,蒸腾速率增加2.25%~4.75%。本试验条件下,土壤热通量对蒸腾速率影响不明显。
3.2 讨 论
温室作物蒸腾作用受诸如植株自身条件、人为因素(环境控制)、室内气象要素等条件的共同影响,本研究基于冬季温室很少通风,室内风速很低甚至为0的特定环境下,采用了根据叶温与周围空气温度差值计算单株植株边界层空气动力学阻力与气孔平均阻力反演模型,并运用了日光温室冬季番茄蒸腾量估算模型。
基于模型模拟蒸腾速率的准确程度与边界层空气动力学阻力、气孔平均阻力的取值有直接关系[2],而二者数值的精确获得很难。风速及温度的变化会产生不同的气流模式类型,若采用“大叶子”假设则与实际作物生产情况存在差异[31];气孔平均阻力的取值不仅与太阳辐射、VPD、空气温度、作物水分等有关[7],且还受植株结构、植株上方气流及土壤条件等影响[14]。本研究采用了与室内气象要素相符合的单株植株边界层空气动力学阻力及反演的气孔平均阻力模型,经检验单株蒸腾速率模拟结果较好。
太阳辐射是作物蒸腾的驱动力,R.de.Graaf等[32]研究表明对于有夜间加温的玻璃温室冬季番茄幼苗,5%的外部太阳总辐射通过作物蒸腾作用转化为潜热,对于生长茂盛的番茄,转化率为37%;汪小旵等[8]、罗卫红等[7]通过对南方现代化温室黄瓜夏季及冬季蒸腾研究表明,到达作物冠层上方的净辐射转化为潜热的比率分别为66%和46%;Meir Teitel[10]研究防虫遮阳网温室内番茄蒸腾发现9∶00—14∶00大约70%~80%的净辐射通过作物蒸腾作用转化为潜热;本研究中,单株植株上方净辐射量的43.5%通过植株蒸腾作用转化为潜热,太阳能转化率介于上述研究结果之间,这也是基质袋培条件下北方寒区冬季日光温室特定气象条件所产生的结果。
本试验中番茄为基质袋培,地面覆盖塑料薄膜,无裸露土壤。土壤热通量变化对逐时蒸腾速率影响仅在白天一段时间略超过5%,其余时间均低于5%,由此可见在本试验条件下土壤热通量对逐时蒸腾速率的影响不明显,此结论可为相似栽培方式提供参考。
由于番茄植株营养生长和生殖生长可同时进行,番茄植株定植后15 d左右第一穗果开花随即结果,此后可通过测量各时期植株叶面积指数和温室内气象要素计算植株蒸腾量,以此为北方寒区日光温室冬季基质袋培番茄全生育期滴灌灌溉量提供参考。
与大型玻璃、塑料温室相比,日光温室内光照、温度、湿度具有空间分布不均匀的特征。本研究选取温室内中间部位的植株进行测量,可能会高估整体蒸腾量。日光温室冬季生产时光照、温度、湿度的管理是非常重要的,其中湿度的管理要由灌溉量决定,灌溉量少会干旱,灌溉量多会使室内湿度上升进而增加植物病虫害发生频率,因此基于P-M方程估算单株番茄蒸腾量可对番茄的水分管理提供参考。
