长期不同施肥潮土活性有机氮库组分与酶活性对外源牛粪的响应
2019-09-12戚瑞敏温延臣赵秉强林治安李志杰
戚瑞敏,温延臣,赵秉强,林治安,李志杰,李 娟
(中国农业科学院农业资源与农业区划研究所/农业农村部植物营养与肥料重点实验室,北京 100081)
氮是地球上最丰富的化学元素和植物生长必需的营养元素之一。氮循环是农业生态系统中最基本的物质循环过程,对农业生态系统的稳定性、生产力及其环境效应具有关键性的作用[1-2]。施肥是影响土壤质量演化及其可持续利用最为深刻的农业措施之一[3],长期施肥不仅能够提高土壤供氮能力,还能稳定土壤氮库,促进土壤“供氮、保氮”,使进入土壤中的氮迅速转化为土壤的活性氮库和稳定氮库[4-5]。土壤活性有机质 (微生物量、可溶性有机质及颗粒有机质) 中的氮组分,即微生物量氮 (MBN)、可溶性有机氮 (DON) 和颗粒有机氮 (PON) 被认为是土壤活性氮库中的重要组分,在土壤氮循环过程中起着非常重要的作用[6-9]。了解长期不同施肥土壤活性有机氮库组分来源及外源碳添加对其参与土壤氮循环的动态转化过程及特征尤为重要。
土壤有机氮库是土壤氮库的主要存在形式,约占土壤氮库的85%,是土壤氮素的主体与作物必需氮素的主要来源[10],主要受土壤质地和类型、种植制度、农艺方式 (耕作、施肥、灌溉等) 及气热条件等环境因素的影响,外界环境因素不同,土壤生态系统有机氮库的动态平衡不同,其各组分的物理、化学性质和对作物的有效性也不同[4,8,11-12]。土壤有机氮库组分是土壤有机氮的重要化学形态,按照提取方法 (物理、化学、生物)、组成成分及活性程度不同,有不同的定义与分级[13]。国内外常用的土壤有机氮的分级是Bremner1965年提出的方法,主要根据土壤有机氮的化学形态组成成分将有机氮分为氨态氮、氨基酸态氮、氨基糖态氮、酸解未知氮和酸不溶氮,且对不同生境条件及农艺措施对土壤有机氮分级特征的影响进行了广泛研究[9,14-16]。近些年,有研究根据物理-化学分组方法将土壤有机氮分为未保护游离态颗粒、物理保护、化学保护及生物化学保护有机氮组分[17-18]并进行了深入研究[19-20]。根据活性程度不同,土壤有机氮库可分为活性有机氮库与惰性有机氮库[5,21-22];而活性有机氮组分中,颗粒有机氮 (PON)、可溶性有机氮 (DON)、轻组有机氮(LFON) 以及微生物量氮 (MBN) 被认为是土壤活性氮库中的重要组成成分[7,23]。国内外对不同生态系统土壤有机氮库单个组分研究较多[22],但仅限于单个组分“数量”或“含量”的研究,而对不同生境条件下土壤活性氮库组分的系统研究较少,对其参与土壤氮循环过程中调控机制的“质”方面的研究更少[15,24]。前人研究表明,土壤可溶性有机氮及提取态有机氮参与了土壤氮矿化过程,且动态转化过程一致,对土壤氮矿化起着重要作用[9,25],农田土壤中,DON占土壤全氮的0.15%~0.19%[26],虽然土壤可溶性氮在土壤全氮中的占比很小,但是却被认为是参与动态变化及生物可利用矿质氮源中最重要的部分[27];有机肥、秸秆还田等措施增加了土壤可溶性氮及提取态有机氮含量[25]。颗粒态有机氮 (PON) 占土壤氮的比例主要在1.5%~10.0%[28],其短期内动态变化能够反映土壤氮循环特性[29-30]。15N标记秸秆示踪研究表明,PON的矿化是农业土壤矿质氮的主要来源,矿化速率受作物秸秆的C/N比及土壤特性等影响[9]。Hurisso等[31]的研究表明,由长期耕作导致的土壤活性氮的退化单靠免耕是不能恢复的,指出需通过外源物添加提高土壤活性有机氮库。
土壤有机氮的转化主要是土壤微生物及其分泌的酶共同作用的结果[32],因此,对土壤酶参与有机氮矿化的生物学机制的深入了解有利于从本质上认识不同土壤有机氮矿化差异的来源。大量研究表明,施肥尤其是施用有机肥能直接或间接地影响土壤酶活性[33-34]。前期研究结果表明,与不施肥相比,施用有机肥能显著增加土壤水解酶[α-葡萄糖苷酶 (AG)、β-葡萄糖苷酶 (BG)、β-木糖苷酶 (BXYL)、纤维二糖水解酶 (CBH) 和磷酸酶 (PHOS) ]活性,却显著降低了土壤酚氧化酶 (phenox) 活性,且施用有机肥对土壤酶活性的影响随有机肥施用量的增加而增加[35]。长期不同施肥土壤中有机氮库和酶活性不同,其对添加外源有机物的响应不同,参与外源有机物分解转化的过程及其调控机制也有所不同。因此,本研究以始建于1986年的长期定位施肥试验为平台,通过室内培养试验研究添加牛粪条件下,不同施肥制度潮土活性有机氮库及酶活性变化特征,揭示不同施肥潮土活性有机氮库对牛粪的响应机制,为提升土壤肥力,维持土壤生态平衡和改善土壤生物学特性提供理论依据,对于实现资源利用及农业可持续发展具有重要的指导意义。
1 材料与方法
1.1 试验设计
供试土壤采自于中国农业科学院德州实验站禹城试验基地的有机无机配比长期定位试验 (建于1986年10月),该试验地位于华北平原东部 (东经116°34′、北纬 36°50′),光热资源丰富,太阳辐射总量5225 MJ/m2,年日照时数2640 h,≥ 0℃积温4951℃,≥ 10℃积温4441℃,年平均气温13.1℃;雨热同期,年平均降雨量570 mm,水面蒸发量2094 mm,无霜期206天;属暖温带半湿润季风气候。试验地的土壤类型为潮土,成土母质为黄河冲积物,耕层质地轻壤 (黏粒含量21.4%,粉粒含量65.6%,砂粒含量3.0%);初始土壤 (0—20 cm) (1986年) 的基本理化性状分别为:有机碳3.93 g/kg,全氮0.51 g/kg,有效氮37.5 mg/kg,速效磷7.5 mg/kg,速效钾73.0 mg/kg,阳离子交换量15.8 cmol/kg,水溶性盐含量0.96 mg/kg,pH值8.56。主要种植制度为冬小麦-夏玉米一年两熟制。试验设6个处理,本研究选取其中的4个处理,分别为:1) 不施肥 (对照,CK);2) 常量有机肥 (OF,有机养分年施用N 375~450 kg/hm2;P、K按照有机肥氮量进行折算);3) 常量化肥 (CF,年施用N 375~450 kg/hm2、P2O5225~300 kg/hm2、K2O 150 kg/hm2);4) 常量有机无机配施 (1/2有机肥 + 1/2化肥,OCF)。常量有机肥与化肥处理的年养分投入量相同,每个处理重复4次,小区之间用0.8 m的水泥挡板隔开,小区面积28 m2(4 m × 7 m)。具体作物品种、肥料性质、施肥措施及小区试验设计详见Li等[36]的研究。
土壤样品于2014年10月玉米收获后取样,每个小区随机选取5~10个点,用土钻 (3 cm) 取0—20 cm的土样混匀,装好用冰盒立即带回实验室并迅速测定土壤样品含水量。将土壤样品分成两部分,一部分风干用于土壤理化性质的测定;另一部分过2 mm筛并存于4℃,用于后续的室内培养试验。其理化性质见表1。
选取对照 (CK) 和三个常量施肥处理 (OF、CF、OCF) 共16个土壤样品进行室内培养试验。称取相当于100 g干土重的鲜土 (培养前将土壤含水量调至田间含水量的60%) 于250 mL培养瓶中分别进行不添加 (M0) 和添加牛粪 (M1) 两个处理的培养试验,同时用不添加土壤的培养瓶作为对照,在25℃恒温培养箱中培养79天,每个土壤样品设2个重复,定期打开培养瓶塞通气,使瓶内气体与大气进行充分交换,同时用称重法调节土壤含水量使其保持田间含水量的60%。在培养第25天及79天时,取出土样保存于4℃下用于土壤全氮及活性有机氮库组分的测定分析,并测定第79天土壤酶活性。
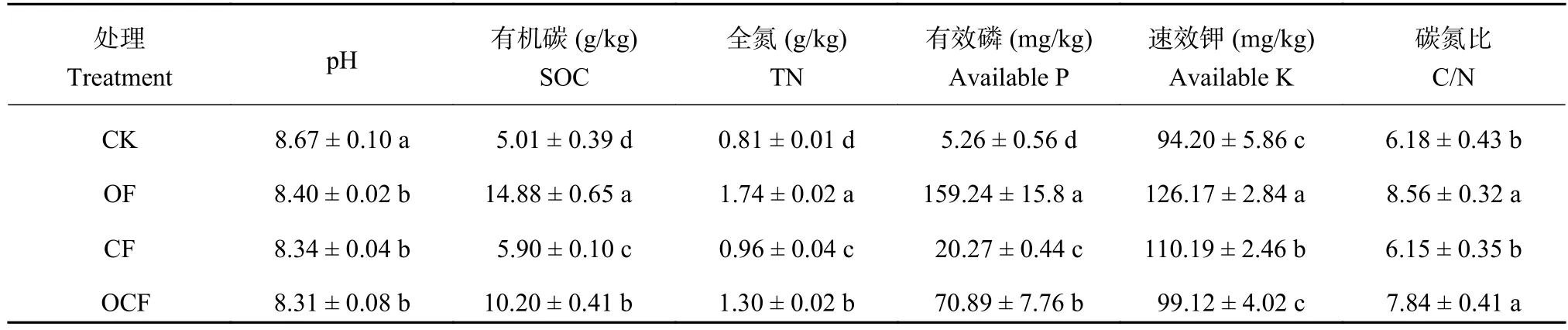
表1 2014年玉米收获后各处理土壤化学性状Table 1 Chemical properties of soils in each treatment after maize harvest in 2014
牛粪选用2014年禹城当地畜牧养殖场腐熟牛粪,主要养分含量为碳 (C) 21.5%、氮 (N) 1.67%、磷(P2O5) 2.68%、钾 (K2O) 1.57%,碳氮比 (C/N) 为12.88,牛粪的施用量按与长期定位施肥中常量化肥施氮量等量折算,为每kg干土施N 0.2 g。
1.2 测定项目与方法
1.2.1 土壤全氮 采用凯氏定氮法。土样经浓硫酸消煮后,再将碱化蒸馏出来的氨用硼酸吸收,以标准酸溶液滴定,求土壤全氮含量。
1.2.2 微生物量氮 (MBN) 土样经氯仿熏蒸-K2SO4浸提后,用TOC/N自动分析仪 (Multi N/C®205 3100,德国耶拿) 测定。称取预培养后的新鲜土壤于小烧杯中,将其置于真空抽滤器中用去乙醇氯仿在25℃的培养箱中避光熏蒸24 h。然后反复抽滤几次以去除氯仿,将熏蒸的土壤样品用0.5 mol/L K2SO4溶液振荡浸提,与此同时以相同的方法提取未熏蒸的土壤,浸提液用TOC/N自动分析仪 (Multi N/C ®205 3100,德国耶拿) 测定。微生物量氮计算公式:BN= EN/KEN。其中,EN为熏蒸和未熏蒸土壤的差值;KEN为转换系数,取值0.45[38-39]。
1.2.3 可溶性有机氮 (DON) 0.5 mol/L K2SO4提取法-TOC/N自动分析仪 (Multi N/C ®205 3100,德国耶拿) 测定。
称取相当于10 g干土的鲜土于125 mL的塑料瓶中,加入50 mL 0.5 mol/L K2SO4,在往复振荡机中振荡 (250 r/min) 提取1 h,然后2500 r/min离心15 min,上清液通过真空泵抽滤过0.45 μm的滤膜后,用TOC/N自动分析仪 (Multi N/C ®205 3100,德国耶拿) 测定其DON含量[40-41]。
1.2.4 颗粒态有机氮 (PON) 采用5 g/L六偏磷酸钠提取法测定。称取过2 mm筛的风干土10 g于125 mL的塑料瓶中,加入30 mL 5 g/L的六偏磷酸钠溶液,在往复振荡机中振荡18 h (90 r/min),分散。分散液置于53 μm的筛上,用蒸馏水冲洗至滤液澄清。将筛上53~2000 μm的土样冲洗至烧杯中,60℃烘干至恒重,计算其占整个土壤样品的百分比,并测定烘干样品中的全氮含量,将烘干样品中的土壤全氮含量换算成整个土壤样品的对应的颗粒态氮组分含量[42]。
1.2.5 土壤酶活性的测定 荧光微孔板酶活性分析法能够在短时间内测定大批量的土壤样品和多种酶活性,近年来被广泛应用于不同生态系统的土壤酶活性研究中[37]。本研究中,测定了参与土壤碳氮磷循环的7种土壤酶活性,其中包括α-葡萄糖苷酶 (α-1,4-glucosidase,AG)、β-葡萄糖苷酶 (β-1,4-glucosidase,BG)、β-木糖苷酶 (β-1,4-xylosidase,BXYL)、纤维二糖水解酶 (cellobiohydrolase,CBH)和磷酸酶 (phosphatase,PHOS) 5种水解酶及过氧化物酶 (peroxidase,perox) 和酚氧化酶 (phenol oxidase,phenox) 2种氧化还原酶。
利用黑色微孔板测定了7种水解酶活性,并根据DeForest[43]的研究方法,对使用的缓冲液稍作调整。具体步骤如下:称取相当于1.0 g干土重的鲜土于200 mL的塑料瓶中,加入125 mL去离子水作为缓冲液,在振荡机上振荡30 min,制成均质泥浆。然后用排枪加200 μL土壤悬浮液于96孔微孔板中(每个样品做8个平行),整个过程需用磁力搅拌器搅匀土壤悬浮液。样品分析孔中加入200 μL土壤悬浮液和50 μL底物溶液;阴性对照孔中加入200 μL缓冲液和50 μL底物溶液;样品对照孔中加入200 μL土壤悬浮液和50 μL缓冲液;标准孔中加入200 μL缓冲液和50 μL标准溶液 (10 mmol/L的4-甲基伞形酮);淬火对照孔中加入200 μL土壤悬浮液和50 μL标准溶液。将微孔板于25℃黑暗条件下培养4小时,随后用酶标仪 (Scientific Fluoroskan Ascent FL,Thermo) 检测荧光值,检测的激发光和发射光波长分别为365 nm和450 nm。土壤酶活性[nmol/(g·h)]用单位时间内每克土样转化底物的nmol数表示。
过氧化物酶、酚氧化酶活性用L-DOPA作为底物在白色微孔板中进行测定[43-44]。空白对照孔中加入250 μL缓冲液;阴性对照孔中加入200 μL缓冲液和50 μL的L-DOPA溶液 (浓度为25 mmol/L);样品对照孔中加入200 μL土壤悬浮液和50 μL缓冲液;样品分析孔中加入200 μL土壤悬浮液和50 μL的L-DOPA溶液。对于过氧化物酶活性的分析,每个孔中均需加入10 μL 0.3%的双氧水。将白色微孔板置于25℃黑暗条件下培养20 h,随后用酶标仪在450 nm的波长下测定吸光度,其活性用μmol/(g·h) 表示。
1.3 数据处理
采用Origin8作图,分析土壤活性有机氮组分的变化特征;用Canoco5进行冗余分析,研究土壤酶活性与活性有机氮库组分之间的关系;采用Excel 2013及SAS 8.0软件进行统计分析,LSD法检验差异显著性 (P < 0.05)。
2 结果与分析
2.1 不同施肥制度土壤活性有机氮库的变化特征
土壤活性有机质中的氮组分作为土壤活性氮库中的重要组成成分,受施肥的影响较大。长期施肥28年后,与长期施化肥 (CF) 及不施肥处理 (CK) 相比,长期施有机肥 (OF) 能显著提高土壤活性有机氮库组分 (MBN、DON和PON) 含量,长期施化肥处理 (CF) 效果不及长期施有机肥 (OF) 及有机无机配施处理 (OCF),OCF中的DON含量最高,CK和OF中MBN的含量差异不显著 (图1),这与本课题组之前报道一致[5]。这说明施肥有利于土壤活性氮的储存,特别是PON的含量,有利于提高土壤的供氮能力。研究表明,长期施肥土壤活性氮库组分(MBN、DON和PON) 与土壤全氮含量之间均呈显著性正相关,且活性有机氮库组分相互之间也显著正相关 (表 2)。
2.2 牛粪对不同施肥土壤有机氮库的影响
由图1可以看出,培养期间,土壤TN及PON含量变化趋势在各处理中均为:OF > OCF > CF >CK (P < 0.05);OF处理中无论添加牛粪与否,TN及PON含量均随着时间的延长呈先降低后逐渐平稳趋势 (图1);在不添加牛粪情况下,除OF处理的PON含量显著下降外 (P < 0.05),CF,OCF和CK处理的PON含量随培养时间的延长变化不显著;与不添加牛粪相比,添加牛粪显著增加了OCF、CF及CK处理中TN及PON含量 (P < 0.05)。
除OCF处理外,其他施肥处理土壤MBN含量随培养时间延长均为先增加再降低的趋势;OCF处理中土壤MBN含量随培养时间延长逐渐降低。不添加牛粪处理的CF和CK处理土壤MBN含量始终显著高于添加牛粪处理 (P < 0.05),但添加牛粪情况下,OF和OCF处理土壤MBN含量在培养第25天均大于不添加牛粪处理,在第79天时,MBN含量均显著低于不添加牛粪处理 (P < 0.05) (图1)。无论添加牛粪与否,各施肥处理土壤DON含量随培养时间延长均呈上升趋势;培养期间,除OF外,其他处理DON含量均为添加牛粪的处理与不添加牛粪处理的相当 (第25天) 或者高于不添加牛粪处理 (第79天),而OF处理的DON含量表现为在第25天,不添加牛粪处理的显著高于添加牛粪处理,而到第79天,添加牛粪的处理又高于不添加牛粪的处理。
连续培养25天,与不添加牛粪相比,添加牛粪显著增加了各处理土壤TN及PON含量,对CK及CF处理的作用效果显著高于OF及OCF处理;除CF处理外,较不添加牛粪相比,添加牛粪显著降低了其他处理的DON含量;相反,添加牛粪显著增加了OF及OCF处理MBN含量,而显著降低了CK及CF处理含量 (图2)。
培养79天后,与不添加牛粪相比,添加牛粪显著增加了各处理土壤TN、DON及PON组分含量,分别提高了5.43%~15.49%、5.83%~69.42%和19.75%~42.29%。图2显示,添加牛粪对CK及长期施用化肥处理 (CF) 的影响比对长期施有机肥处理(OF、OCF) 的影响明显。但添加牛粪显著降低了土壤MBN含量,降低幅度为16.91%~62.10%,且对长期施有机肥处理 (OF、OCF) 的影响显著大于对CK及CF的影响。

图 1 好气培养下添加牛粪对长期不同施肥土壤全氮及活性有机氮库组分含量的影响Fig. 1 Dynamic changes of soil total N and labile organic nitrogen fractions in soils under different fertilizer patterns during incubation
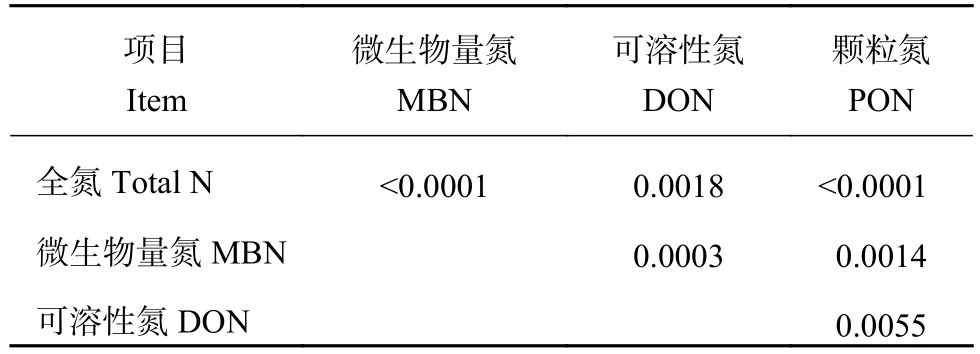
表2 长期不同施肥土壤活性有机氮库组分之间及其与土壤全氮含量间相关性分析 (P值)Table 2 Correlation analysis between soil total nitrogen and labile organic nitrogen fractions (P value)
2.3 牛粪对不同施肥制度下土壤酶活性的影响
表3分析表明,施肥、添加牛粪及其交互作用对土壤酶活性具有显著影响 (P< 0.05)。
为明确不同施肥处理下土壤酶活性的变化规律,计算了添加与不添加牛粪处理的土壤酶活性的百分比差异 (图3),无论添加牛粪与否,三个施肥处理土壤过氧化物酶和酚氧化酶活性显著低于对照土壤,水解酶活性却呈现不同的变化趋势。不添加牛粪培养,CF处理土壤β-木糖苷酶 (BXYL) 和β-纤维素酶 (CBH) 分别提高了209%和181%。添加牛粪培养,OF处理土壤β-葡萄糖苷酶 (BG) 和β-纤维素酶(CBH) 分别提高了201%和308%。
2.4 添加牛粪土壤有机氮库与酶活性之间的相关性分析
将不同施肥制度土壤的TN、MBN、DON、PON和SOC/TN (C/N) 作为自变量 (解释变量),土壤酶活性作为因变量 (响应变量),分别进行了添加和不添加牛粪条件下的冗余分析 (redundancy analysis,RDA)。在不添加牛粪时 (图4),解释变量可解释模型的48.30%,轴1和轴2分别占36.95%和9.08%。施肥处理 (CF、OF、OCF) 的组群与CK处理显著分离,分别在轴的正反两方向上,并且不同施肥处理也显著分开。预选 (forward-selection) 结果表明,DON是影响土壤酶活性变化的重要影响因素,可以解释总变量的34.40% (P< 0.01),这可能是造成不同施肥处理之间分异的主要原因。
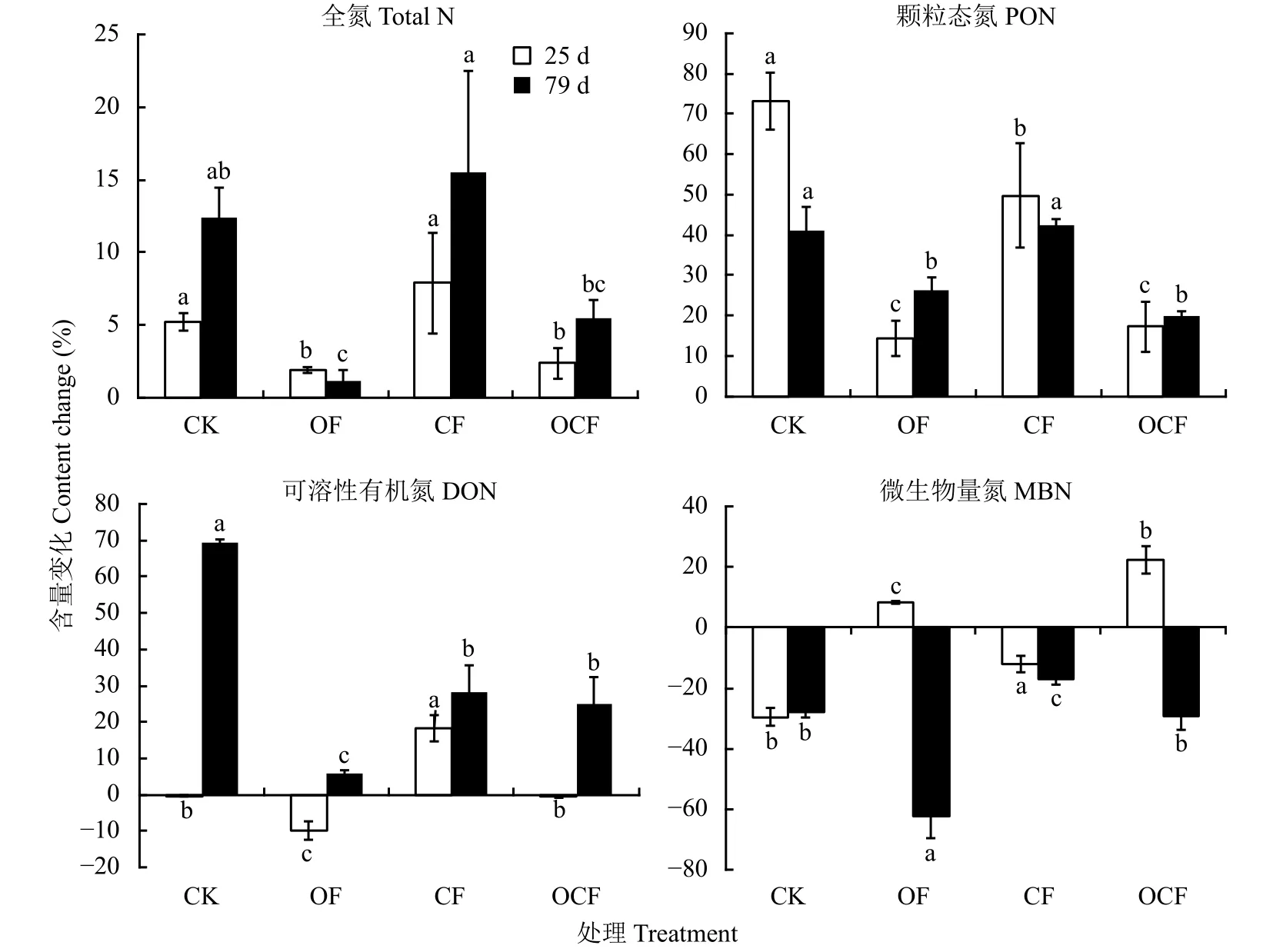
图 2 不同施肥处理土壤添加牛粪培养后全氮及有机氮组分含量的相对变化Fig. 2 Relative change of total N and organic N fractions affected by cattle manure addition in soils under different fertilization patterns
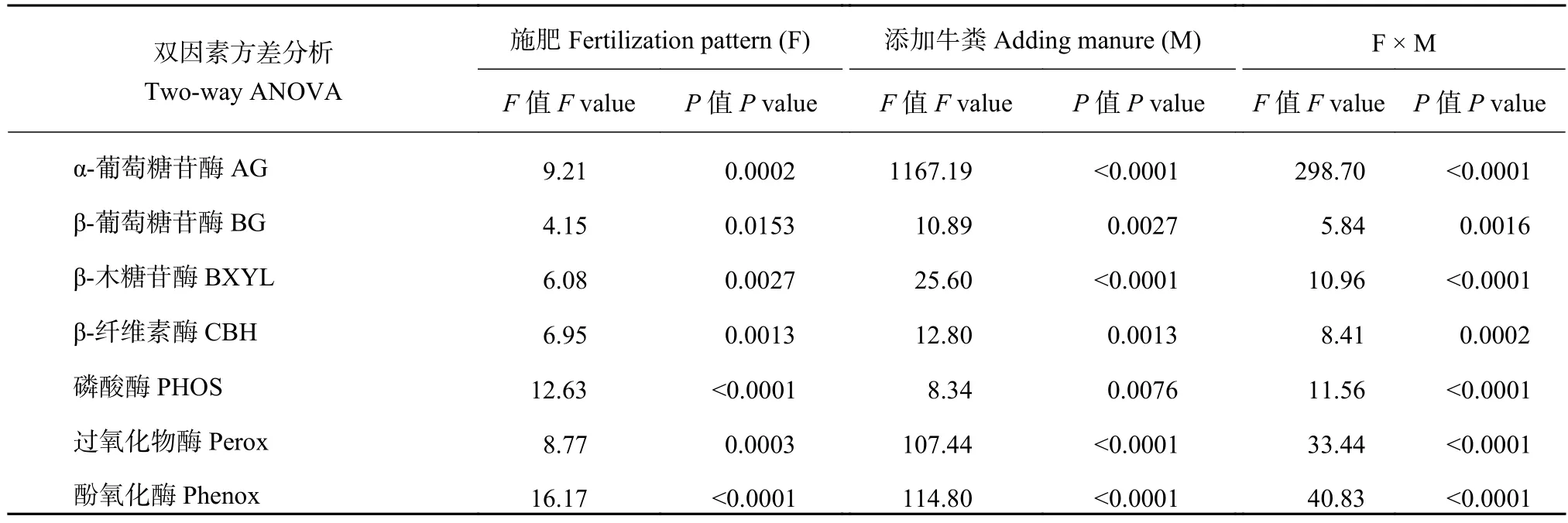
表3 施肥、添加牛粪对土壤酶活性变化的影响 (培养第79天)Table 3 Effect of fertilization pattern and manure addition on soil enzyme activities (Incubating 79 days)
在添加牛粪的RDA模型中 (图4),解释变量占总变量的80.50%,轴1和2的特征根分别解释了变异的56.85%和18.89%。OF、OCF处理与和CK、CF处理分离显著,分别在轴的正反两个方向上。OF与OCF之间以及CK和CF处理之间均有明显的分离。预选结果显示TN和DON是添加牛粪时较好的解释变量因子,能分别解释土壤酶活性变异的56.48% (P< 0.01) 和 18.28% (P< 0.01)。
整体来看,预选模型表明,在土壤全氮和活性有机氮库组分这些环境因子中,添加与不添加牛粪下土壤酶活性的关键驱动因子不同,不添加牛粪条件下可溶性有机氮DON为主要影响因子,添加后其关键影响因子为全氮TN和可溶性有机氮DON。
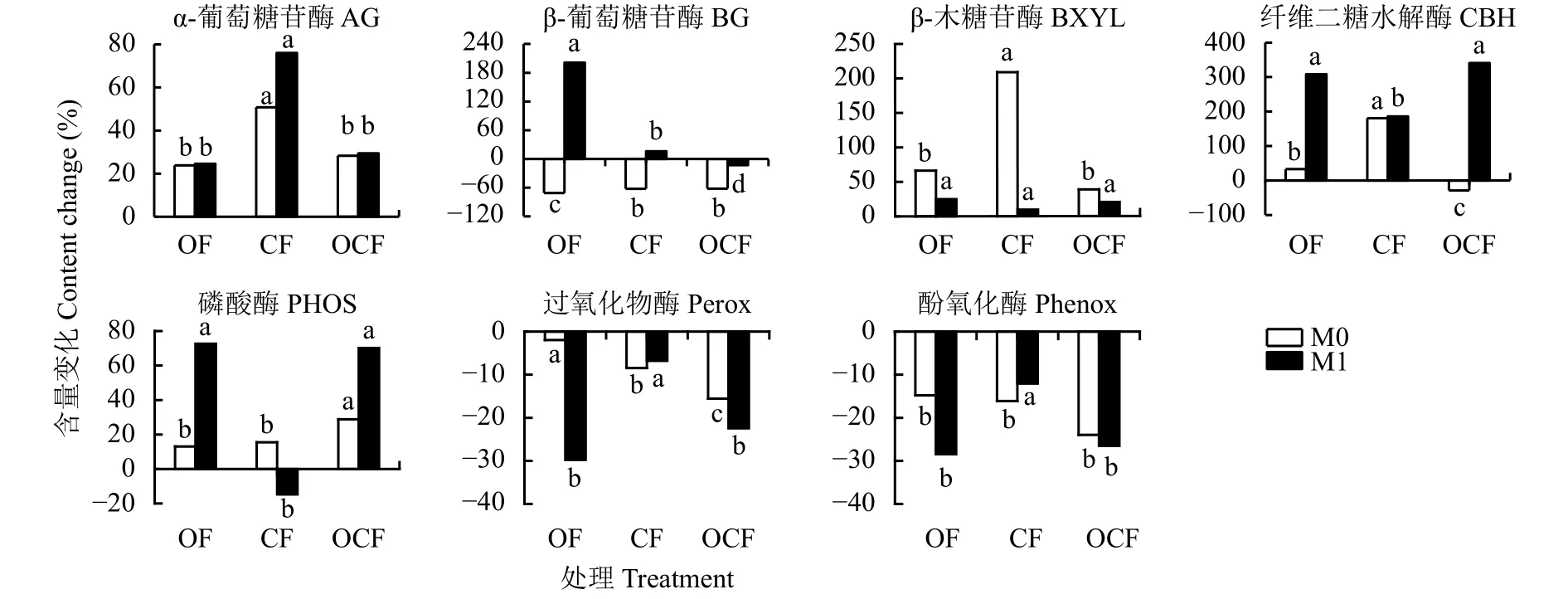
图 3 添加牛粪对各施肥处理土壤酶活性的影响Fig. 3 Effect of cattle manure addition on the enzyme activities of soils from different fertilization patterns

图 4 不同施肥制度土壤酶活性和全氮及有机氮库组分之间的冗余分析Fig. 4 Redundancy analysis of soil enzyme activities constrained by TN and organic nitrogen fractions in soils from different fertilization regimes after 79 days of incubation
3 讨论
土壤可溶性有机氮被用作土壤供氮能力的评价指标及施肥等农艺措施对土壤变化影响的生物指示指标[45]。探讨土壤全氮中随时间变化的活性部分,如土壤微生物量氮,能较好地反映由于固定-矿化过程而改变养分有效性动力的土壤质量和生产能力[46-47],还能较好地估计由于管理措施而引起的氮素矿化潜力变化[48-49]。颗粒态有机氮 (PON) 被认为是处于活性和慢性氮组分之间的中间体,也会随着土壤管理措施的改变作出迅速的响应[48]。
我国区域广阔,各地分布了多个长期定位实验,涵盖了全国不同的土壤类型、气热条件及种植制度。目前为止,已有不少关于不同施肥类型及水平对土壤有机氮库影响的研究工作,可以总结为:长期施肥均能显著增加土壤微生物量氮[50-53]、耕层颗粒有机氮含量及储量[54]。黄威等[8]以湖南3个长期定位试验点为研究对象,指出长期单施化肥、单施有机肥及有机无机配施均能提高红壤活性有机氮含量,且随有机肥施用量的增加而增加;土壤活性氮与全氮的积累速率呈极显著相关。不同种植制度下,北京褐潮土区长期撂荒土壤微生物量氮含量显著高于农田土壤[50],且在农田土壤中,长期轮作的土壤微生物量氮含量要高于长期连作的;韩晓日等[55]指出施有机肥对黑土土壤微生物量氮贡献大,施化肥对土壤微生物量氮的贡献较小,土壤氮是微生物量氮的主要来源,有机无机配施介于二者之间。本课题组以往研究[5]表明,长期施用有机肥显著提高了黄淮海潮土土壤全氮、土壤微生物量氮、可溶性有机氮和颗粒有机氮含量,且土壤全氮及其活性氮组分的含量随有机肥或化肥施用量的增加而增加。颗粒态氮占土壤全氮的比例最高,为6.6%~18.6%;其次为微生物量氮,为6.8%~8.8%;可溶性有机氮占比最低。这表明,施肥措施不同,对土壤有机氮库组分含量影响及肥料在有机氮库组分中的分配不同[13]。本研究中,微生物量氮、可溶性有机氮和颗粒有机氮与土壤全氮表现出一致的趋势,均随着有机肥施入量的增加而增加,与前人结果相似[50]。有机氮积累过程中,有机肥主要进入土壤颗粒态氮库[5],而化肥可提高土壤微生物量氮在土壤氮库中的百分比[55]。
本研究表明,培养时间及外源有机物料对长期不同施肥土壤活性有机氮库组分的影响也差异显著,影响程度取决于氮源和碳源 (能源) 的供应强度。添加新鲜的有机物料后,土壤碳源充足,激发了土壤微生物的大量生长,生物量氮增加,但当新鲜有机物料被消耗到一定程度后,微生物能利用的易分解有机碳源已耗尽,很快转入利用较难分解的有机碳,生物量氮下降,呈现周期性的变化 (图1)[56]。除土壤微生物量氮外,其他土壤活性氮组分普遍呈现随培养时间延长而逐渐增加趋势,说明土壤微生物在分解有机物料的同时,也分解土壤有机质,转化为PON,释放出DON,从而增加了土壤全氮含量。
土壤肥力不同,有机物料对土壤活性有机氮库含量的影响也不相同。本研究中,培养结束后,添加牛粪对长期施用有机肥土壤的影响效果不及其对长期施化肥及不施肥处理的影响。这与李世清等[14]研究的结果一致,他们认为土壤肥力不同,有机物料对土壤微生物量氮的影响效果不同,有机物料在低肥力土壤上的效果更加突出。胡玮等[57]关于不同碳氮比有机物料对土壤肥力的影响研究表明,碳氮比为12.5的有机物料配比更有利于提高低肥力土壤微生物量氮的含量,这与我们的研究结果一致。本研究结果显示,添加牛粪后,长期不施肥与施化肥处理土壤微生物量氮含量显著低于相同处理不添加牛粪的土壤微生物量氮含量,说明在一定的条件下,牛粪限制了土壤氮向微生物量氮的转化,且刺激了土壤微生物量氮向其他氮库的迁移和转化 (图1)。这与韩晓日等[55]研究结果一致,指出单施有机肥降低了土壤微生物量氮来自土壤氮的百分比,这可能与添加牛粪提高了土壤C/N有关。
土壤酶作为土壤生物活性及土壤肥力的重要组成部分,在土壤物质循环和能量的转化中起着重要的催化作用,其活性可快速反映施肥对土壤肥力和土壤质量的影响[58-59]。本研究表明,长期施肥对土壤酶活性影响很大。对于土壤α-葡萄糖苷酶、β-木糖苷酶BXYL、β-纤维素酶CBH和磷酸酶PHOS总体趋势是施肥的高于不施肥的。施肥之所以能提高土壤酶活性这是因为施肥可促进作物根系代谢,使根系分泌物增多,微生物繁殖加快,从而有利于提高土壤酶活性[60]。对长期施肥土壤酶活性进行meta分析表明,施肥提高了土壤中与有机质分解相关酶(BG和NAG) 的活性,但对与氮循环相关的亮氨酸氨基肽酶 (LAP) 活性无显著影响。原因主要为,施有机肥或化肥直接或间接增加了碳源,促进微生物活动,加速土壤碳循环,提高了土壤中与有机质分解相关酶 (BG和NAG) 的活性[61]。但是,长期施肥却降低了土壤酚氧化酶和过氧化物酶的活性。这与孙瑞莲等[62]报道类似,但与高瑞等[60]报道相反。这一结果可能与土壤类型、气候条件、种植制度等不同有关。
有研究表明,可溶性有机态氮对碱性磷酸酶和蔗糖酶活性有显著影响[63-64],且土壤氮元素的富集减少了过氧化物酶基因的表达量[65],与我们的研究结果,即土壤全氮和可溶性有机氮与土壤酶活性有极强的相关性一致。这说明土壤酶活性的变化与土壤养分的变化之间有直接的关系。土壤酶参与土壤中复杂的生物化学转化过程,研究结果中各酶活性之间极强的相关性说明土壤中进行酶促反应的各土壤酶之间存在相互刺激机制[66-67]。土壤可溶性养分作为土壤现实供肥能力的指示指标,与土壤生物活动及酶活性关系密切[65],这也佐证了我们的研究中可溶性有机氮是调控土壤酶活性关键驱动因子的结果。王涛等[64]研究的退耕刺槐林土壤养分与酶活性关系的结果,表明土壤养分中全氮、总有机碳、可溶性有机氮对酶活性响应最大,这与我们的研究结果一致。
4 结论
长期不同施肥模式显著影响土壤全氮和有机氮库对外源有机物料的响应。添加牛粪显著增加了长期不同施肥土壤全氮、可溶性有机氮和颗粒有机氮含量,却显著降低了土壤微生物量氮含量;其中,
长期施有机肥与有机无机配施处理对外源牛粪的添加响应显著。另外,添加牛粪显著增加了长期施有机肥处理土壤中的β-葡萄糖苷酶 (BG) 和β-纤维素酶(CBH) 活性,显著降低了长期施有机肥及有机无机配施处理过氧化物酶 (perox) 和酚氧化酶 (phenox) 的活性。无论添加牛粪与否,不同施肥制度土壤酶活性之间差异显著;不添加牛粪情况下,土壤酶活性的驱动因子为可溶性有机氮,添加牛粪情况下,可溶性有机氮和全氮成为其关键驱动因子。因此,长期施有机肥及有机无机配施可提高作物对外源有机养分的吸收利用,提高土壤氮库存量,有利于氮素的长期储存。
