梭梭同化枝及其叶苞状虫瘿的转录组差异表达分析
2019-09-10满玲娟张桦姚正培宗兴风李志强
满玲娟 张桦 姚正培 宗兴风 李志强


摘要:【目的】分析梭梭同化枝及其葉苞状虫瘿的转录组差异,为揭示梭梭叶苞状虫瘿形成的分子机制及梭梭害虫防治提供理论参考。【方法】以梭梭同化枝及其叶苞状虫瘿为材料,利用Illumina HiSeq 2000高通量测序平台对其进行转录组测序及差异表达基因分析,并通过测定可溶性糖和叶绿素含量验证分析结果。【结果】转录组测序共获得159147条Unigenes,平均长度为551 bp,N50为752 bp,在Swissport、GO、Pfam、NR、NT、KOG/COG和KEGG等数据库均被注释的Unigenes有8521条(5.35%),至少在1个数据库注释的Unigenes有66714条(41.91%),共有49066条Unigenes(30.83%)与物种匹配成功。以q value<0.005且|log2FoldChange|>1为条件,共筛选出3444条差异表达的Unigenes,其中,有1299条上调表达,2145条下调表达。与梭梭同化枝相比,梭梭叶苞状虫瘿上调表达差异基因富集在代谢过程、有机物代谢过程、催化活性、碳代谢作用和核糖体代谢等通路,下调表达基因富集在细胞、细胞组成、胞内细胞成分和细胞器、嘧啶代谢、细胞周期、DNA复制、光合作用及苯丙烷和类黄酮生物合成等过程。不同发育期梭梭叶苞状虫瘿中的可溶性糖含量高于梭梭同化枝,而叶绿素含量均低于梭梭同化枝,进一步验证梭梭叶苞状虫瘿的糖代谢相关基因上调表达而光合作用相关基因下调表达的转录组分析结果。【结论】梭梭叶苞状虫瘿中核糖体数量增加,翻译过程加快,代谢水平提高,糖代谢产物积累,但其光合作用降低,伸长与生长减慢,推测其通过吸收利用同化枝的养分以满足自身需求。
关键词: 梭梭;叶苞状虫瘿;转录组分析;可溶性糖含量;叶绿素含量
中图分类号: S792.99 文献标志码: A 文章编号:2095-1191(2019)08-1657-08
Differential expression analysis of transcriptome of assimilating branches and leafy-bracted galls of Haloxylon ammodendron
MAN Ling-juan, ZHANG Hua*, YAO Zheng-pei, ZONG Xing-feng, LI Zhi-qiang
(College of Agronomy, Xinjiang Agricultural University, Urumqi 830052, China)
Abstract:【Objective】The transcriptome difference of the leafy-bracted gall and assimilating branch of Haloxylon ammodendron was analyzed to provided a theoretical reference for exploring the molecular mechanism of leafy-bracted gall formation and the pest control of H. ammodendron. 【Method】The experimental materials were the assimilating branches of H. ammodendron and the leafy-bracted gall. The transcriptome was sequenced using Illumina HiSeq 2000 high-throughput sequencing platform,and differential expression gene was analyzed. The soluble sugar and chlorophyll contents of them were measured to confirm the results of transcriptome analysis. 【Result】A total of 159147 Unigenes were obtained with average length of 551 bp. Their N50 was 752 bp. In Swissport, GO, Pfam, NR, NT, KOG/COG and KEGG,8521 Unigenes(5.35%) were successfully annotated in all databases, 66714 Unigenes(41.91%) were annotated in at least one database,and 49066 Unigenes(30.83%) were successfully matched to species in total. Using q value<0.005 and |log2FoldChange|>1 as screening condition,3444 Unigenes were differentially expressed,among which 1299 were up-regulated and 2145 were down-regulated. Compared with the assimilating branches of H. ammodendron,the up-regulated genes in gall were mainly enriched in metabolic processes,organic substance metabolic process,catalytic activities,carbohydrate metabolic process and ribosome pathway,while the down-regulated genes were enriched in cells,cell component,intracellular component and organelle,pyrimidine metabolism,cell cycle,DNA replication,photosynthesis,phenylpropanoid biosynthesis and flavonoid biosynthesis. The soluble sugar content in galls of different developmental stages was higher than that of assimilating branches,while the chlorophyll content was lower than that of assimilating branches. The results further confirmed that the genes related to sugar metabolism were up-regulated,but the genes related to photosynthesis were down-regulated in transcriptome analysis. 【Conclusion】The number of ribosomes in the leafy-bractedgall of H. ammodendron increases,the translation process accelerates,the metabolic level increases,and the glycometabolic metabolites accumulates,but its photosynthesis decreases,its elongation and growth slow down. It is inferred that the gall can supply its own needs by absorbing and utilizing the nutrients of assimilating branches.
Key words: Haloxylon ammodendron; leafy-bracted gall; transcriptome analysis; soluble sugar content; chlorophyll content
0 引言
【研究意义】梭梭(Haloxylon ammodendron)为藜科(Chenopodiaceae)梭梭属(Haloxylon)植物,其叶片退化为鳞状叶,由同化枝进行光合作用,生长在沙丘、盐渍土及砾质戈壁等地,能耐受高温、干旱、盐碱、风蚀的恶劣环境,具有防风固沙的生态保护作用(俞阗,2012;Long et al.,2014)。木虱(Psyllidae)可严重危害梭梭生长,其在梭梭同化枝上引起的虫瘿外形类似叶苞状,被称为梭梭叶苞状虫瘿(姜梦辉等,2017)。虫瘿是因昆虫、病菌等刺激诱导寄主植物产生的一种畸形瘤状物或突起(邵淑霞等,2012;Carneiro et al.,2017),其形成和发育是植物与昆虫共同作用的结果,明确其形成过程对探讨生物间协同进化和减少昆虫对寄主的危害十分重要(王光钺等,2010;申洁等,2016)。因此,利用转录组测序分析梭梭同化枝及其虫瘿的基因表达差异有助于深入探究虫瘿生长发育及形成的分子机制,对分析木虱与梭梭的进化关系及开展梭梭林虫害防治具有重要意义。【前人研究进展】通过转录组测序可获取特定生长状况下某组织或细胞的转录本信息,从而分析基因的表达情况,现已发展成为深入挖掘未知基因的重要手段(刘永杰,2016;Hrdlickova et al.,2017;唐玉娟等,2018)。目前,利用该技术开展了大量关于虫瘿的研究工作。如桃金娘叶片及其虫瘿的转录组测序结果表明,虫瘿形成与生长素响应基因相关(Bailey et al.,2015);白蜡虫真蛹的转录组测序结果表明,白蜡虫蛹期生理活动与蛋白降解和Wnt信号通路等相关(于淑惠等,2016);长叶木姜子及其杯状瘿的转录组测序结果表明,杯状瘿内能量或营养相关基因的改变促使其光合作用降低,可溶性糖含量升高(Shih et al.,2018);榆树叶片及其虫瘿的转录组测序结果表明,虫瘿的形成与糖代谢、次生代谢、植物激素信号转导等代谢通路相关(李轩和黄智鸿,2019);葡萄叶片及其虫瘿的转录组测序结果表明,虫瘿的形成与形成层分生组织、生长素信号转导等途径相关(Schultz et al.,2019)。关于梭梭叶苞状虫瘿的相关研究主要集中在虫瘿的形态和生态学方面,李粉莲等(2012)研究发现,梭梭叶苞状虫瘿内有异色胖木虱、梭梭胖木虱、矮胖木虱和显赫胖木虱4种木虱;姜梦辉等(2017)通过对梭梭同化枝及其叶苞状虫瘿进行野外和显微结构观察,将虫瘿的生长发育分为生长期、形成期和衰亡期3个阶段,且虫瘿表皮细胞的叶绿体较梭梭同化枝少。【本研究切入点】至今,鲜见有关梭梭叶苞状虫瘿形成机制及转录组差异表达分析的研究报道。【拟解决的关键问题】以梭梭同化枝及其叶苞状虫瘿为材料,对其进行转录组测序及差异表达基因分析,并通过测定可溶性糖和叶绿素含量验证分析结果,旨在了解虫瘿的发育机理及对梭梭生长的影响,为揭示梭梭叶苞状虫瘿形成的分子机制及害虫防治提供理论参考。
1 材料与方法
1. 1 试验材料
在新疆古尔班通古特沙漠南缘选取生长健康且高度基本一致的梭梭植株,采集梭梭同化枝及其生长期和形成期的叶苞状虫瘿为试验材料(图1),液氮速冻后置于超低温冰箱保存备用。TRIzol植物总RNA提取试剂盒购自北京全式金生物技术有限公司,DNase I(RNase-free)试剂盒购自Thermo公司,其他生化试剂均购自新疆皓博晟达生物科技有限公司。主要仪器设备:DYY-6B型电泳仪(北京六一仪器厂)、Agilent Bioanalyzer 2100生物芯片分析仪(Agilent,美国)、NanoDrop 1000分光光度计(Thermo,美国)和722型可见分光光度计(上海菁华科技仪器有限公司)。
1. 2 转录组测序
利用TRIzol植物总RNA提取试剂盒分别提取梭梭同化枝及其叶苞状虫瘿的总RNA,用1.0%琼脂糖凝胶电泳检测其质量。经DNase I(RNase-free)试剂盒纯化,使用Agilent Bioanalyzer 2100生物芯片分析仪和NanoDrop 1000分光光度计分别检测总RNA的纯度及浓度。分别将检测合格的同化枝总RNA溶液及生长期和形成期的叶苞状虫瘿总RNA等体积混合液送至北京诺禾致源生物信息科技有限公司进行转录组测序。测序得到的原始序列raw reads经过滤除杂得到高质量clean reads,再利用Trinity对clean reads进行拼接和组装,最终获得的Unigenes作为后续分析的参考序列(Grabherr et al.,2011)。
1. 3 转录组功能注释
获得的Unigenes与NR、NT、Pfam、KOG/COG、Swissprot、GO和KEGG等数据库比对注释。其中,根据Nr数据库注释结果可得到与Unigenes比对的物种分布图,并基于NR和Pfam数据库注释信息,使用Blast2GO对Unigenes进行GO功能注释及分类(Conesa et al.,2005)。根據KEGG数据库将参与各代谢通路的Unigenes进行分类。
1. 4 基因表达水平分析及差异表达基因筛选
参考Li和Dewey(2011)的方法,利用RSEM(bowtie2为默认参数)将每个样品的clean reads与参考序列进行比对分析,统计与参考序列匹配成功的clean reads,获得各样品比对到每个基因的read count数目,对其进行FPKM(每百万fragments中来自某一基因每千碱基长度的fragments数目)转换,当FPKM>0.3时判定为基因表达。
使用TMM将read count数据标准化处理后,以DEGseq进行差异表达分析:筛选出阈值为q value (校正后的p value)<0.005且|log2FoldChange|>1的基因,当基因的log2FoldChange>0,则认为该基因表达上调,反之,则认为该基因表达下调(Anders and Huber,2010)。使用GOseq方法对获得的差异表达基因(Differentially expressed genes,DEGs)进行GO富集分析(Young et al.,2010);同时采用KOBAS 2.0进行KEGG富集分析(Kanehisa et al.,2008)。
1. 5 可溶性糖和叶绿素含量的测定
为证实转录组测序结果的准确性,分别测定梭梭同化枝及其叶苞状虫瘿中的可溶性糖和叶绿素含量。采集梭梭同化枝及不同发育期的叶苞状虫瘿(图2),采用蒽酮比色法测定其可溶性糖含量(张桦,2014),采用乙醇丙酮混合液浸提法测定其叶绿素含量(徐芬芬等,2010),各样品均设3个重复。
2 结果与分析
2. 1 转录组测序结果
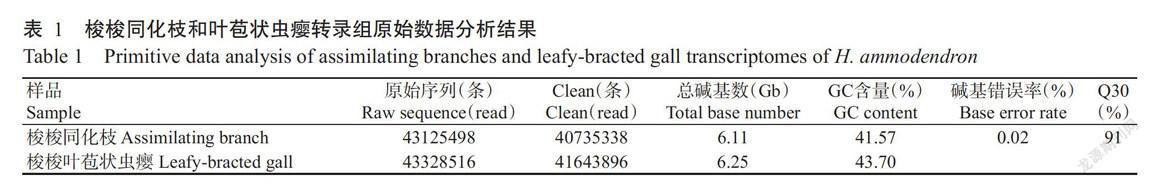
梭梭同化枝及其叶苞状虫瘿的转录组测序结果如表1所示,GC含量分别为41.57%和43.70%,Q30为91%,表明测序数据质量较好,可用于后续分析。高质量clean reads经拼接组装后获得转录本序列,取每条基因中最长的转录本作为Unigene,最终获得159147条Unigenes,平均长度为551 bp,N50为752 bp,其中序列长度最短为201 bp,最长为13228 bp,200~500 bp的序列有113933条,大于2 kb的序列有6296条。
2. 2 Unigenes的功能注释结果
统计在各数据库注释的Unigenens数目,结果表明,159147条Unigenes中,在NR数据库注释的Unigenes有49128条(30.86%),在Swissport、GO、Pfam、Nt、KOG/COG和KEGG等数据库注释的Unigenens分别有43404条(27.27%)、43396条(27.26%)、42925条(26.97%)、29768条(18.70%)、27121条(17.04%)和20561条(12.91%),在以上7个数据库均被注释的Unigenes有8521条(5.35%),至少在1个数据库注释的Unigenes有66714条(41.91%)。
与Nr数据库中已知物种序列进行比对分析,发现共有49066条Unigenes(30.83%)与物种匹配成功,其中匹配序列数量最多的为甜菜(Beta vulgaris)25025条(51.00%),其次为林烟草(Nicotiana sylvestris)2368条(4.80%)、绒毛状烟草(N. tomentosiformis)2263条(4.60%)、大麦(Hordeum vulgare)974条(2.00%)和蓝隐藻(Guillardia theta)701条(1.40%),与其他物种匹配序列共计17735条(36.10%),有110081条(69.17%)未匹配成功。
2. 3 差异表达基因的GO富集分析结果
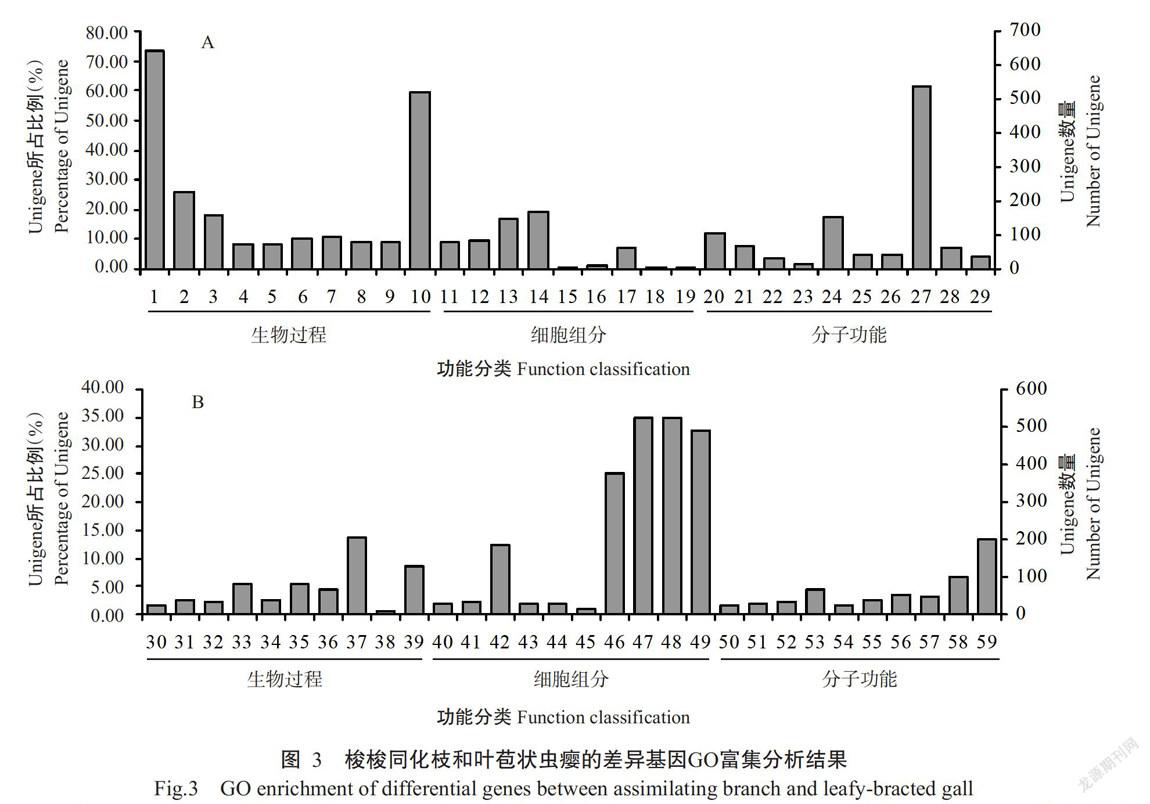
根据Nr注释信息,对Unigenes进行GO分类,结果发现共有43396条Unigenens注释到生物过程(Biological process)、分子功能(Molecular function)和细胞组分(Cellular component)中的56个亚类。以q value<0.005且|log2FoldChange|>1为条件,共筛选出3444条差异表达的Unigenes,其中1299条上调表达、2145条下调表达。差异表达基因的GO富集分析结果显示,有3819条Unigenes注释到生物过程的20个亚类,560条Unigenes注释到细胞组分的9个亚类,1448条Unigenes注释到分子功能的20个亚类。
将GO富集分类中前10个富集项(q value<0.05)在图3中依次列出。与梭梭同化枝相比,梭梭叶苞状虫瘿上调表达基因在生物过程类型中主要富集于代谢过程、有机物代谢过程、有机氮复合代谢和氧化还原等过程;在细胞组分类型中主要富集于细胞质、核糖体、核糖核蛋白复合体等;在分子功能类型中主要富集于催化活性和氧化活性等功能方面,说明梭梭叶苞状虫瘿较其同化枝的代谢水平高。与梭梭同化枝相比,梭梭叶苞状虫瘿下调表达基因在生物过程类型中主要富集于细胞成分组织或生物的发生、核糖体及核蛋白体发生、细胞或亚细胞成分的运动、光合作用和微管等过程;在细胞组分类型中主要富集于细胞、细胞器、细胞成分和微管等组分;在分子功能类型中主要富集于DNA结合、结构分子活性、核糖体结构成分和微管蛋白结合等功能方面,说明梭梭叶苞状虫瘿光合作用下降,细胞的伸长与生长受到影响。
2. 4 差异表达基因的KEGG通路分析结果
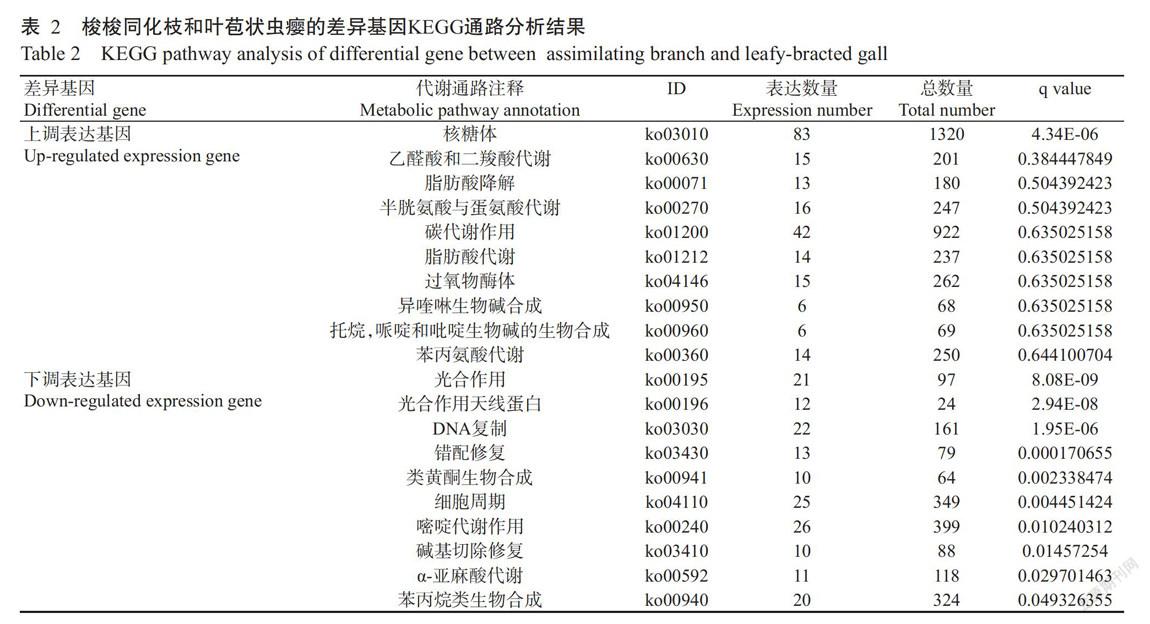
差异表达基因主要参与五类KEGG通路,分别为细胞过程、环境信息处理、遗传信息处理、代谢和有机系统。其中,在细胞过程通路中,参与运输与分解代谢的相关基因最多,为1107条;在环境信息处理通路中,参与信号转导和膜转运的相关基因较多,分别为475和149条;在遗传信息处理通路中,参与翻译的相关基因最多,为2627条,其次为参与蛋白质折叠、分类和降解过程、转录及复制与修复的相关基因,分别为1718、1041和392条;在代谢通路中,参与碳水化合物代谢的相关基因最多,为2005条,其次为参与新陈代谢、氨基酸代谢和能量代谢的相關基因分别为1500、1337和1121条;在有机系统通路中,参与环境适应的相关基因最多,为639条。
差异表达基因参与的代谢通路在表2中按照q value依次列出。与梭梭同化枝相比,梭梭叶苞状虫瘿中上调表达基因主要富集在核糖体代谢通路,其次为核糖体、乙醛酸和二羧酸代谢、脂肪酸降解、碳代谢作用和脂肪酸代谢等通路,推测这些基因上调表达会引起梭梭叶苞状虫瘿糖代谢产物的积累。下调表达基因主要富集在光合作用、光合作用天线蛋白、DNA复制、细胞周期、嘧啶代谢作用、苯丙烷类生物合成和类黄酮生物合成等通路,表明与梭梭同化枝比较,梭梭叶苞状虫瘿的光合效率降低,同时细胞分裂和木质素合成受影响。
2. 5 梭梭同化枝和叶苞状虫瘿的可溶性糖和叶绿素含量测定结果
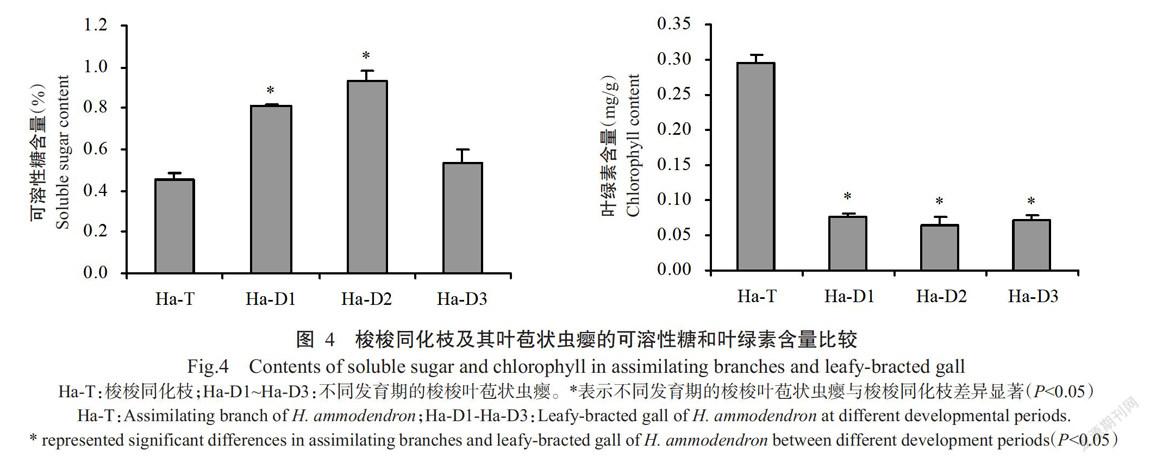
分别对梭梭同化枝及其叶苞状虫瘿进行可溶性糖和叶绿素含量测定,结果如图4所示。随着梭梭叶苞状虫瘿的生长,可溶性糖含量呈先升高后降低的变化趋势,其中Ha-D1和Ha-D2叶苞状虫瘿的可溶性糖含量显著高于同化枝(P<0.05,下同),但Ha-D1、Ha-D2和Ha-D3叶苞状虫瘿的叶绿素含量均显著低于同化枝。可见,与梭梭同化枝相比,梭梭叶苞状虫瘿的可溶性糖含量较高,叶绿素含量较低,进一步验证梭梭叶苞状虫瘿的糖代谢相关基因上调表达而光合作用相关基因下调表达的转录组分析结果。
3 讨论
本研究的梭梭同化枝和叶苞状虫瘿转录组测序及功能注释结果显示,159147条Unigenes中,在NR、NT、Pfam、KOG/COG、Swissprot、GO和KEGG等数据库中均被注释的Unigenens有8521条(5.35%),至少在1个数据库注释的Unigenens有66714条(41.91%),但仅有49066条Unigenes(30.83%)与物种匹配,表明梭梭同化枝和叶苞状虫瘿中仍有大量未知的Unigenes,这些序列可能编码新蛋白。Bailey等(2015)对桃金娘叶片及其虫瘿进行转录组差异分析,结果表明,在正常桃金娘叶片中存在未表达却因虫瘿形成而重新启动的基因。同样,本研究发现只在梭梭叶苞状虫瘿中表达的Unigenes有530条,推测这些Unigenes是与叶苞状虫瘿形成有关的特异表达序列,为今后挖掘相关基因提供了参考。
虫瘿通过消耗较高的能量以维持其旺盛的生长和新陈代谢,导致寄主植物组织的结构和功能发生改变,光合作用减弱(Carneiro et al.,2017)。可溶性糖为植物生长提供能量并构成结构物质,在植物生长过程中发挥重要作用(Ma et al.,2017)。前人研究发现,荔枝和无患子的虫瘿中可溶性糖含量比正常叶片高,说明可溶性糖是虫瘿代谢、发育及昆虫生长的营养物质(Huang et al.,2015;Oliveira et al.,2017)。本研究的GO富集分析和KEGG通路分析结果显示,梭梭叶苞状虫瘿上调表达差异基因主要富集在代谢过程、有机物代谢过程、催化活性、碳水化合物代谢和核糖体代谢等通路,说明梭梭叶苞状虫瘿中核糖体数量增加,翻译过程加快,代谢水平提高,糖代谢产物积累,为梭梭叶苞状虫瘿及若虫的生长提供能量和条件,与不同发育阶段梭梭叶苞状虫瘿的可溶性糖含量均高于梭梭同化枝的结果相互印证。此外,本研究发现不同发育阶段梭梭叶苞状虫瘿的叶绿素含量均低于梭梭同化枝,与梭梭叶苞状虫瘿下调表达基因富集在光合作用和光合作用天线蛋白的结果相互印证,表明叶苞状虫瘿的光合作用降低。前人研究也发现,樱桃和无患子叶片的虫瘿内叶绿素含量较低,光合作用下降(Larson,1998;Oliveira et al.,2017),与本研究结果基本一致。梭梭叶苞状虫瘿的光合作用降低,代谢活性升高,推测其通过吸收利用梭梭同化枝的养分以满足自身需求。
微管在植物生长发育中参与细胞内运输、细胞运动及分裂过程,同时构成细胞骨架,维持细胞形态,且参与合成植物中的纤维素和木质素等(邱鸿和于荣,2009;饶国栋和张建国,2013),而苯丙烷类代谢产物可进一步转化成类黄酮、木质素等,在植物木质化和自我保護过程中发挥重要作用(Grace and Logan,2000;孟雪娇,2011;王星等,2017)。本研究的GO富集分析和KEGG通路分析结果显示,下调表达基因富集在微管及其过程、微管运动及其活性、微管结合、微管蛋白结合、结构分子活性、苯丙烷和类黄酮生物合成、细胞周期和DNA复制等过程。这些基因与细胞分裂、运动和伸长生长相关,其表达量降低不利于营养物质的长距离运输、细胞伸长及形态维持,可能是造成梭梭叶苞状虫瘿较短小的主要原因,而苯丙烷、类黄酮等次生代谢产物合成相关基因的表达量降低,可能是导致梭梭叶苞状虫瘿质地较软且木质化程度下降的主要因素。但关于叶苞状虫瘿是如何影响梭梭同化枝正常生长而形成现有形态、如何改变次生代谢而利于幼虫生长均有待进一步研究。
4 结论
梭梭叶苞状虫瘿中核糖体数量增加,翻译过程加快,代谢水平提高,糖代谢产物积累,但其光合作用降低,伸长生长减慢,推测其通过吸收利用同化枝的养分以满足自身的需求。
参考文献:
姜梦辉,姚正培,任燕萍,姜子焱,谷月好,王波,张桦. 2017. 梭梭叶苞状虫瘿结构特征和生长发育研究[J]. 西北植物学报,37(9):1749-1755. [Jiang M H,Yao Z P,Ren Y P,Jiang Z Y,Gu Y H,Wang B,Zhang H. 2017. Study on the structure characteristics and growth and development of leafy galls in Haloxylon ammodendron[J]. Acta Bota-nica Boreali-Occidentalia Sinica,37(9):1749-1755.]
李粉莲,吴雪海,王佩玲,梁虎军,王鑫,王利军,张建萍. 2012. 梭梭木虱发生规律及其影响因子[J]. 生态学报,32(8):2311-2319. [Li F L,Wu X H,Wang P L,Liang H J,Wang X,Wang L J,Zhang J P. 2012. The occurrence regularity of psyllid in Haloxylon spp and its influencing factors[J]. Acta Ecologica Sinica,32(8):2311-2319.]
李轩,黄智鸿. 2019. 榆树叶片形成虫瘿过程中转录组学分析[J]. 基因组学与应用生物学,38(2):737-746. [Li X,Huang Z H. 2019. Transcriptome analysis in the process of insect gall formation of Elm leaves[J]. Genomics and Applied Biology,38(2):737-746.]
刘永杰. 2016. 玉米抗禾谷镰刀菌的转录组分析[D]. 北京:中国农业大学. [Liu Y J. 2016. Transcriptome analysis of maize resistance to Fusariiim graminearum[D]. Beijing:China Agricultural University.]
孟雪娇. 2011. 黄瓜苯丙烷类代谢关键酶活性及基因表达的研究[D]. 哈尔滨:哈尔滨师范大学. [Meng X J. 2011. The study on activities of cucumber phenylpropanoid metabolism’s key enzymes and gene expression[D]. Harbin:Harbin Normal University.]
邱鸿,于荣. 2009. 微管和微丝的相互作用[J]. 细胞生物学杂志,31(4):476-480. [Qiu H,Yu R. 2009. The interactions of microtubules and microfilaments[J]. Chinese Journal of Cell Biology,31(4):476-480.]
饶国栋,张建国. 2013. 植物微管蛋白基因研究进展[J]. 世界林业研究,26(3):17-20. [Rao G D,Zhang J G. 2013. Advances of studies on plant tubulin gene[J]. World Fores-try Research,26(3):17-20.]
邵淑霞,阮桢媛,杨子祥,陈晓鸣. 2012. 虫瘿—昆虫与植物互作的奇特产物[J]. 环境昆虫学报,34(3):363-369. [Shao S X,Ruan Z Y,Yang Z X,Chen X M. 2012. Insect galls:The peculiar product of the interaction between plants and in-sects[J]. Journal of Environmental Entomo-logy,34(3):363-369.]
申洁,董颖颖,王义平. 2016. 虫瘿与其生物群落及寄主植物间的关系[J]. 环境昆虫学报,38(2):445-450. [Shen J,Dong Y Y,Wang Y P. 2016. The relationships between insects galls and its biological community and their host plants[J]. Journal of Environmental Entomology,38(2):445-450.]
唐玉娟,黄国弟,罗世杏,周俊岸,莫永龙,李日旺,赵英,张宇,宋恩亮,宁琳. 2018. 芒果2个不同花芽分化时期转录组分析[J]. 南方农业学报,49(7):1257-1264. [Tang Y J,Huang G D,Luo S X,Zhou J A,Mo Y L,Li R W,Zhao Y,Zhang Y,Song E L,Ning L. 2018. Transcriptome of Mangifera indica L. in two different flower bud differentiation stages[J]. Journal of Southern Agriculture,49(7):1257-1264.]
王光鉞,王义平,吴鸿. 2010. 虫瘿与致瘿昆虫[J]. 昆虫知识,47(2):419-424. [Wang G Y,Wang Y P,Wu H. 2010. Gall and gall-former insects[J]. Chinese Bulletin of Entomology,47(2):419-424.]
王星,罗双霞,于萍,罗蕾,赵建军,王彦华,申书兴,陈雪平. 2017. 茄科蔬菜苯丙烷类代谢及相关酶基因研究进展[J]. 园艺学报,44(9):1738-1748. [Wang X,Luo S X,Yu P,Luo L,Zhao J J,Wang Y H,Shen S X,Chen X P. 2017. Advances in phenylaprapanoid metabolism and its enzyme genes in Solanaceae vegetables[J]. Acta Horticulturae Sinica,44(9):1738-1748.]
徐芬芬,叶利民,徐卫红,郑静萍. 2010. 小白菜叶绿素含量的测定方法比较[J]. 北方园艺,(23):32-34. [Xu F F,Ye L M,Xu W H,Zheng J P. 2010. Comparison of methods of chlorophyll extraction in Chinese cabbage[J]. Northern Horticulture,(23):32-34.]
于淑惠,亓倩,孙涛,王雪庆,杨璞,冯颖. 2016. 白蜡虫雄虫真蛹转录组分析[J]. 林业科学研究,29(3):413 417. [Yu S H,Qi Q,Sun T,Wang X Q,Yang P,Feng Y. 2016. Transcriptome analysis of male white wax scale pupae[J]. Fore-st Research,29(3):413-417.]
俞阗. 2012. 荒漠植物梭梭对地表高温胁迫的响应及“花后生殖休眠”现象的研究[D]. 南京:南京农业大学. [Yu T. 2012. Study on H. ammondendron for response to land surface temperature stress and phenomenon of reproductive dormancy after flower[D]. Nanjing:Nanjing Agricultural University.]
張桦. 2014. 生物化学实验指导[M]. 北京:中国农业出版社. [Zhang H. 2014. Biochemical experimental guidance[M]. Beijing:China Agriculture Press.]
Anders S,Huber W. 2010. Differential expression analysis for sequence count data[J]. Genome Biology,11(10):R106.
Bailey S,Percy D M,Hefer C A,Cronk Q C. 2015. The transcriptional landscape of insect galls:Psyllid(Hemiptera) gall formation in Hawaiian Metrosideros polymorpha(Myrtaceae)[J]. BMC Genomics,16:943. doi:https://doi.org/10.1186/s12864-015-2109-9.
Carneiro R G S,Isaias R M S,Moreira A S F P,Oliveira D C. 2017. Reacquisition of new meristematic sites determines the development of a new organ,the cecidomyiidae gall on Copaifera langsdorffii desf.(Fabaceae)[J]. Frontiers in Plant Science,8:1622.
Conesa A,Gotz S,Garcia-Gomez J M,Terol J,Talon M,Robles M. 2005. Blast2GO:A universal tool for annotation,visualization and analysis in functional genomics research[J]. Bioinformatics,21(18):3674-3676.
Grabherr M G,Haas B J,Yassour M,Levin J Z,Thompson D A,Amit I,Adiconis X,Fan L,Raychowdhury R,Zeng Q D,Chen Z,Mauceli E,Hacohen N,Gnirke A,Rhind N,Birren B W,Nusbaum C,Lindblad-Toh K,Friedman N,Regev A. 2011. Full-length transcriptome assembly from RNA-Seq data without a reference genome[J]. Nature Biotechnol,29(7):644-652.
Grace S C,Logan B A. 2000. Energy dissipation and radical scavenging by the plant phenylpropanoid pathway[J]. Phi-losophical Transactions of the Royal Society of London,Series B,Biological Sciences,355(1402):1499-1510.
Hrdlickova R,Toloue M,Tian B. 2017. RNA-Seq methods for transcriptome analysis[J]. Wiley Interdiscip Rev RNA,8(1):10. doi:10.1002/wrna.1364.
Huang M Y,Huang W D,Chou H M,Chen C C,Chen P J,Chang Y T,Yang C M. 2015. Structural,biochemical,and physiological characterization of photosynthesis in leaf-derived cup-shaped galls on Litsea acuminata[J].
BMC Plant Biology,15:61. doi:https://doi.org/10.1186/
s12870015-0446-0.
Kanehisa M,Araki M,Goto S,Hattori1 M,Hirakawa M,Itoh M,Katayama T,Kawashima S,Okuda S,Tokimatsu T and Yamanishi Y. 2008. KEGG for linking genomes to life and the environment[J]. Nucleic Acids Research,36:480-484.
Larson K C. 1998. The impact of two gall-forming arthropods on the photosynthetic rates of their hosts[J]. Oecologia,115(1):161-166.
Li B,Dewey C N. 2011. RSEM:Accurate transcript quantification from RNA-Seq data with or without a reference genome[J]. BMC Bioinformatics,12:323.
Long Y,Zhang J W,Tian X J,Wu S S,Zhang Q,Zhang J P,Dang Z H,Wu X. 2014. De novo assembly of the desert tree Haloxylon ammodendron(C. A. Mey.) based on RNA-Seq data provides insight into drought response,gene discovery and marker identification[J]. BMC Genomics,15(1):1111-1122.
Ma Q J,Sun M H,Lu J,Liu Y J,Hu D G,Hao Y J. 2017. Transcription factor AREB2 is involved in soluble sugar accumulation by activating sugar transporter and amylase genes[J]. Plant Physiology,174(4):2348-2362.
Oliveira D C,Moreira A,Isaias R,Martini V,Rezende U C. 2017. Sink status and photosynthetic rate of the leaflet galls induced by Bystracoccus mataybae(Eriococcidae) on Matayba guianensis(Sapindaceae)[J]. Frontiers in Plant Science,8:1249. doi:10.3389/fpls.2017.01249.
Schultz J C,Edger P P,Body M,Appel H. M. 2019. A galling insect activates plant reproductive programs during gall development[J]. Scientific Reports,9(1):1833.
Shih T H,Lin S H,Huang M Y,Sun C W,Yang C M. 2018. Transcriptome profile of cup-shaped galls in Litsea acumi-nata leaves[J]. PLoS One,13(10):e0205265.
Young M D,Wakefield M J,Smyth G K,Oshlack A. 2010. Gene ontology analysis for RNA-seq:Accounting for selection bias[J]. Genome Biology,11:R14. doi:https://doi.org/10.1186/gb-2010-11-2-r14.
(責任编辑 陈 燕)
