绣球菌多糖及其体外免疫活性研究
2019-09-10张迪王宏雨肖冬来林衍铨
张迪 王宏雨 肖冬来 林衍铨


摘要:[目的]分析绣球菌水溶性多糖的结构与免疫活性。[方法]采用热水提取法从广叶绣球菌Sparassislatifolla的真空冷冻干燥品中提取得到绣球菌水溶性多糖SCG-D,然后通过DEAE Sepharose FastFlow离子交换层析对其进行分离,获得SCG-N和SCG-A两个多糖组分,并对SCG-A组分进行HPSEC、单糖组成分析、红外光谱、H-NMR和C-NMR核磁共振的结构分析。通过大鼠脾淋巴细胞体外刺激活性测试比较了SCG-D及其组分SCG-A、SCG-N的体外免疫活性差异。[结果]绣球菌水溶性多糖SCG-D具有促进鼠脾淋巴细胞增殖的活性,其酸性多糖组分SCG-A促进作用最强,其100ug.mL72h处理组的增殖率为32.7%,而SCG-N多糖组分的活性较SCG-A低,其100ug·mL72h处理组增殖率为11.66%。结构分析结果表明SCG-A的重均分子量为4.30×10Da,其主链结构为α-1,4-D-葡聚糖,主要由葡萄糖和半乳糖构成,并且还含有一定量的1-6分支结构。[结论]绣球菌冻干品的水溶性多糖及其組分具有促进大鼠脾淋巴细胞体外增殖的免疫活性,其中活性最强的酸性多糖组分SCG-A是一种具有1-6分支结构的α-l,4-葡聚糖。
关键词:广叶绣球菌;多糖;脾淋巴细胞;免疫
中图分类号:S646.9文献标志码:A 文章编号:1008-0384(2019)09-1093-07
0引言
(研究意义)广叶绣球菌Sparassis latifolia,属担子菌亚门Basidiomycotina、异隔担子菌纲Heteroba-sidiomycetes、无(非)褶菌目ApHyllopHorales、绣球菌科Sparassidacea、绣球菌属Jparassis Fr.。绣球菌具有很高的营养保健价值,绣球菌中含有大量的活性多糖,能够提高人体免疫力及机体造血功能,对某些肿瘤也有一定的预防和抑制作用。(前人研究进展)白辰等的试验结果表明绣球菌子实体冻干品的可溶性多糖保留率显著高于热风干燥品。本课题组在前期研究中发现绣球菌子实体冻干品提取的可溶性多糖和子实体烘干品中提取的可溶性多糖分子量构成也有显著不同,烘干品可溶性多糖的主要成分在20KDa左右,而冻干品可溶性多糖的分子量主要分布在400KDa左右。脾脏是人体最大的免疫器官,是淋巴细胞汇集的部位,而淋巴细胞是机体的免疫活性细胞,其激活、分化、增殖在免疫应答过程中起着重要的作用,现有研究结果表明多糖可以与淋巴细胞表面多糖受体相结合,从而影响淋巴细胞的功能,进而调节机体的免疫功能,测定药物对脾淋巴细胞的影响是快速筛选免疫功能成分的有效手段。(本研究切入点)在之前的研究中所述的具有免疫调节活性的绣球菌B-葡聚糖是一种由强碱溶液提取的大分子量葡聚糖,其分子量在200万Da以上,主要结构是带有6分支结构的β-1,3-葡聚糖,难溶于水,低浓度溶液呈凝胶状。而对于绣球菌子实体中的水溶性多糖的一级结构及其生物学活性尚缺乏系统的研究。因此,本研究以前期发现的分子量为400KDa左右的绣球菌可溶性多糖组分为研究对象,通过色谱和波普对其一级结构进行初步分析,并通过大鼠初代脾淋巴细胞体外刺激实验对其免疫调节活性进行检测。(拟解决的关键问题)通过研究明确绣球菌可溶性多糖的主要结构及其体外免疫调节活性,以期探明绣球菌可溶性多糖的生物学活性基础,填补相关研究工作的空白。
1材料与方法
1.1供试材料
绣球菌冻干粉购于福建天益菌业有限公司。
1.2试剂与仪器设备
试剂:Dextran系列葡聚糖(Pharmcia公司);细胞培养瓶(美国FALCON);RPMI-1640培养基(美国GIBCO);FBS(美国ExCellBiologyFBS500);24孔细胞培养板(美国Coming Incorporated);96孔板(美国Coming Incorporated);其余试剂均为国产分析纯、色谱纯;试验用水为超纯水。
仪器:UM 5000蒸发光检测仪(北京通微分析仪器有限公司);SP8800色谱泵(美国物理光谱公司);TSKgel GMPxL凝胶色谱柱(7.8mm×300mm,日本东曹);Waters 2695液相色谱仪(美国WATERS);Waters2998PDA检测器(美国WATERS);ODS-SP色谱柱(150×4.6mm,5um,日本岛津);IX51倒置显微镜(日本OLYMPUS);超净工作台(苏州净化);C0培养箱(日本SANYO);Nicolet-6700红外光谱仪(美国Thermo);AVANCE400核磁共振波谱仪(德国BRUKER);Centrifuge 5804-R离心机(德国艾本德);EPOCH2TC酶标仪(美国BioTek);真空冷冻干燥机(北京博医康)。
1.3试验方法
1.3.1绣球菌多糖提取与分离多糖提取:采用热水提取法,准确称取绣球菌冻干品粉末50g,料液比1:15,100℃热水水浴搅拌提取2h,提取液4000r·min离心15min取上清,上清液浓缩至原体积的1/2,加入4倍体积95%的乙醇醇沉过夜,8000r·min离心10min取沉淀置于真空干燥箱中去除残余乙醇,即得绣球菌粗多糖。
采用三氯乙酸法脱蛋白:绣球菌粗多糖用蒸馏水复溶后,8000r·min离心10min取上清液,用蒸馏水溶解配制成质量浓度为15mg·mL的溶液,加入样品体积的1/10的40%TCA溶液,混匀后静置30min,8000r·min离心10min,取上清。重复3次后,取上清液加入4倍体积乙醇进行醇沉,过夜后4500r·min离心5min取沉淀,用丙酮洗涤沉淀2次,真空干燥后,得到绣球菌冻干品多糖SCG-D。
绣球菌多糖的DEAE Sepharose Fast Flow柱分离:绣球菌冻干品多糖SCG-D溶液(20mg·mL)10mL上样于DEAESepharoseFastFlow柱(2.6cm×25cm)进行分离,流速:1mL·min,先用纯水洗脱,而后用0.25mol·L的NaCl溶液洗脱。以无水乙醇在680nm检测混浊度跟踪多糖,纯水洗脱部分冻干获得多糖组分SCG-N,NaCl溶液洗脱部分用透析袋流水透析48h脱盐后冻干获得多糖组分SCN-A。
1.3.2绣球菌多糖样品的分子量分布测定 多糖分子量分布测定采用高效凝胶渗透色谱法。
色谱条件:流动相为0.05mol·L乙酸铵;流速0.6mL·min;柱温35℃;进样量10uL;色谱柱TSKgel GMPxL(7.5mm×300mm)。ELSD检测器:载气为空气;载气压力4.5bar;漂移管温度60℃;数据分析系统为Chrommanger5.8GPC专用版。
标准品和待测样品的制备:多糖样品和葡聚糖对照品用0.05mol·L乙酸铵溶液溶解为2mg·mL,用于HPSEC分析。
以重均分子量为2000000、1000000、500000、70000、40000、20000Da的葡聚糖样品为标准品,以保留时间对分子量的对数作线性回归。
1.3.3绣球菌SCG-A多糖組分的单糖组成分析 采用反相高效液相色谱(RP-HPLC)结合单糖PMP衍生检测绣球菌SCG-A多糖中的单糖组成及摩尔比。
1.3.4绣球菌SCG-A多糖组分的红外光谱分析 分别称取2mg多糖样品与适量的KBr粉末在白炽灯下干燥研磨均匀,压片机压片,红外光谱仪在400-4000cm区间内扫描透射光谱进行红外分析。
1.3.5绣球菌SCG-A多糖组分的核磁共振分析 20mg多糖样品溶于重水0.5mL,在400Mz核磁共振仪测定,氢谱(300K)时以HDOδ4.69为内标。
1.3.6多糖样品的体外鼠脾淋巴细胞刺激试验 通过MTT法检测SCG-D、SCG-A、SCG-N对大鼠初代脾淋巴细胞体外增值刺激活性。
脾脏淋巴细胞制备:取大鼠无菌操作取出脾脏,用冷PBS漂洗后将脾脏组织剪碎,冷PBS漂洗后加入0.125%胰酶+0.05%Ⅱ型胶原酶进行消化;消化完成后过滤,滤液通过梯度离心收集淋巴细胞,收集的细胞经PBS清洗后于RPMI1640培养基+10%胎牛血清培养。
MTT法检测细胞增殖:将脾淋巴细胞消化、调整浓度为5×10个·mL的细胞悬液备用,在96孔板中每孔加入100uL细胞悬液;然后置于37℃、5%CO环境中培养;24h后每孔加入100uL相应含药培养基,再于37℃、5%CO环境中培养72h;然后进行MTT染色,测定490mm的OD值,计算各组别增殖率,每个处理设3个重复,并设阴性对照组。
细胞增殖率=(处理组OD-对照组OD)/对照组OD×100%
2结果与分析
2.1绣球菌多糖的分离与分子量分布测定
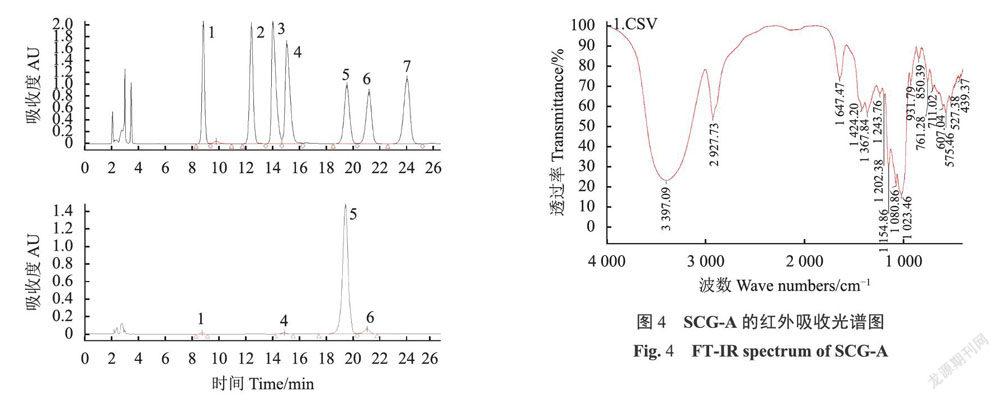
绣球菌水溶性多糖SCG-D通过离子交换树脂分离为不被凝胶柱吸附的SCG-N和被阴离子凝胶柱吸附的酸性多糖SCG-A两部分(图1),分别对其进行HPSEC检测。
标准曲线的回归方程为:Log(MW)=-0.4066tR+10.9773,MW为重均分子量,tR为保留时间(min),相关系数为r=0.9941。从图2可以看出,SCG-A在色谱图上呈现1个明显色谱峰,该成分重均分子量
2.2绣球菌SCG-A多糖组分的单糖组成、红外光谱与核磁共振分析
按文献所述方法,在相同色谱条件下,得到混标单糖PMP衍生物的液相色谱图和SCG-A多糖水解产物PMP衍生物的色谱图(图3),结果显示多糖SCG-A主要由葡萄糖和半乳糖构成,并含有少量的甘露糖和半乳糖醛酸,其中葡萄糖与半乳糖物质量比为22.13:1。
如图4所示,SCG-A的吸收谱带在3397.09cm处存在强且宽的-OH吸收峰;2927.73cm附近的肩峰为饱和C-H伸缩振动的信号,中等强度;1647.47cm处为酰胺羰基吸收峰,表明样品中可能有一定量的糖结合蛋白;1154.86cm,1080.86cm,1023.46cm等3个峰为吡喃糖环特征吸收峰;光谱在890cm附近无吸收峰、而在850cm有一吸收峰,表明其糖苷键构型为α型。综上,SCG-A的主要结构可能为α-D-葡聚糖。
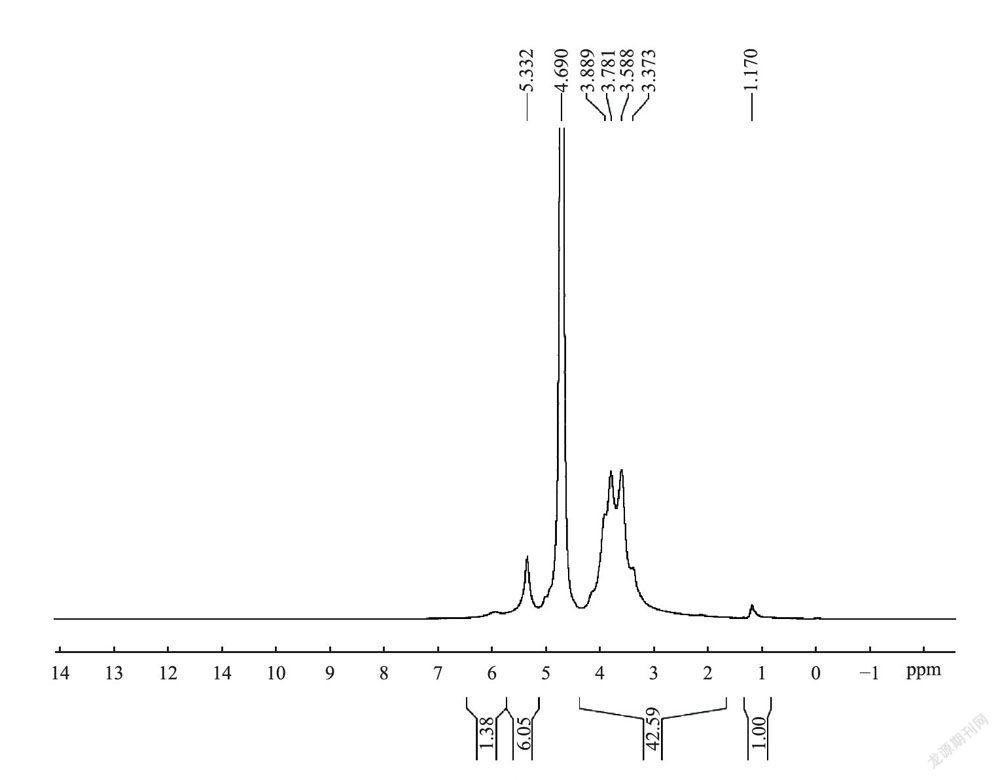
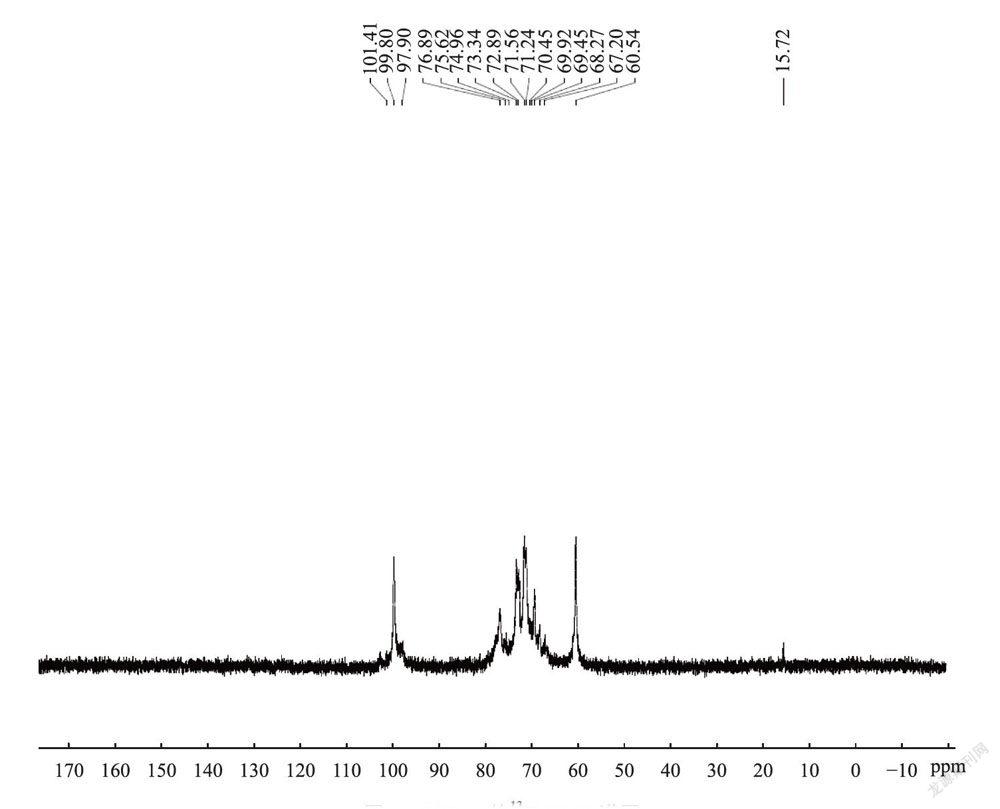
图5和图6分别为多糖组分SCG-A的H-NMR和C_N1VIR谱。在H谱中55.332处为异头氢的共振峰,由于其化学位移大于5.0,因此这些葡萄糖残基均为α型吡喃糖,这与红外光谱分析结论一致。C-NMR谱在δ95-105区域观察到3个端基碳信号,表明其糖链重复单位中应至少有3种单糖,这与单糖分析数据相符,δ99.8处强信号(<100)表明其主链结构苷键为α-D型;在76.89处为C-4发生取代的化学位移信号,δ60.54的强信号表明存在大量未发生取代的C-6,说明糖链主要以(1-4)连接为主;同时,在δ68.34的化学位移信号表明存在有发生取代的C-6,可见糖链中可能还存在(1-6)分支结构。结合单糖组成分析数据和红外光谱数据可知SCG-A为一种以α型1,4糖苷键连接的吡喃型葡萄糖为主链的葡聚糖,同时其可能还含有一定量(1-6)分支结构。
2.3绣球菌多糖组分对大鼠初代脾淋巴细胞的刺激作用
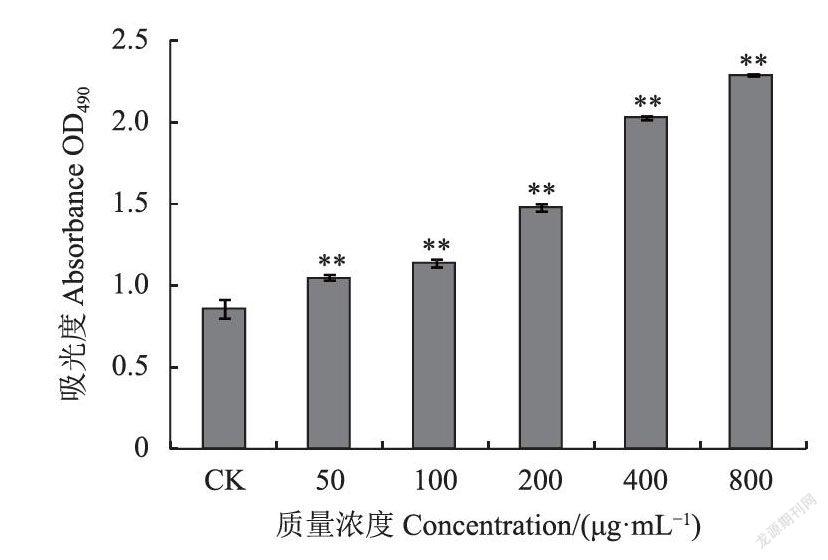
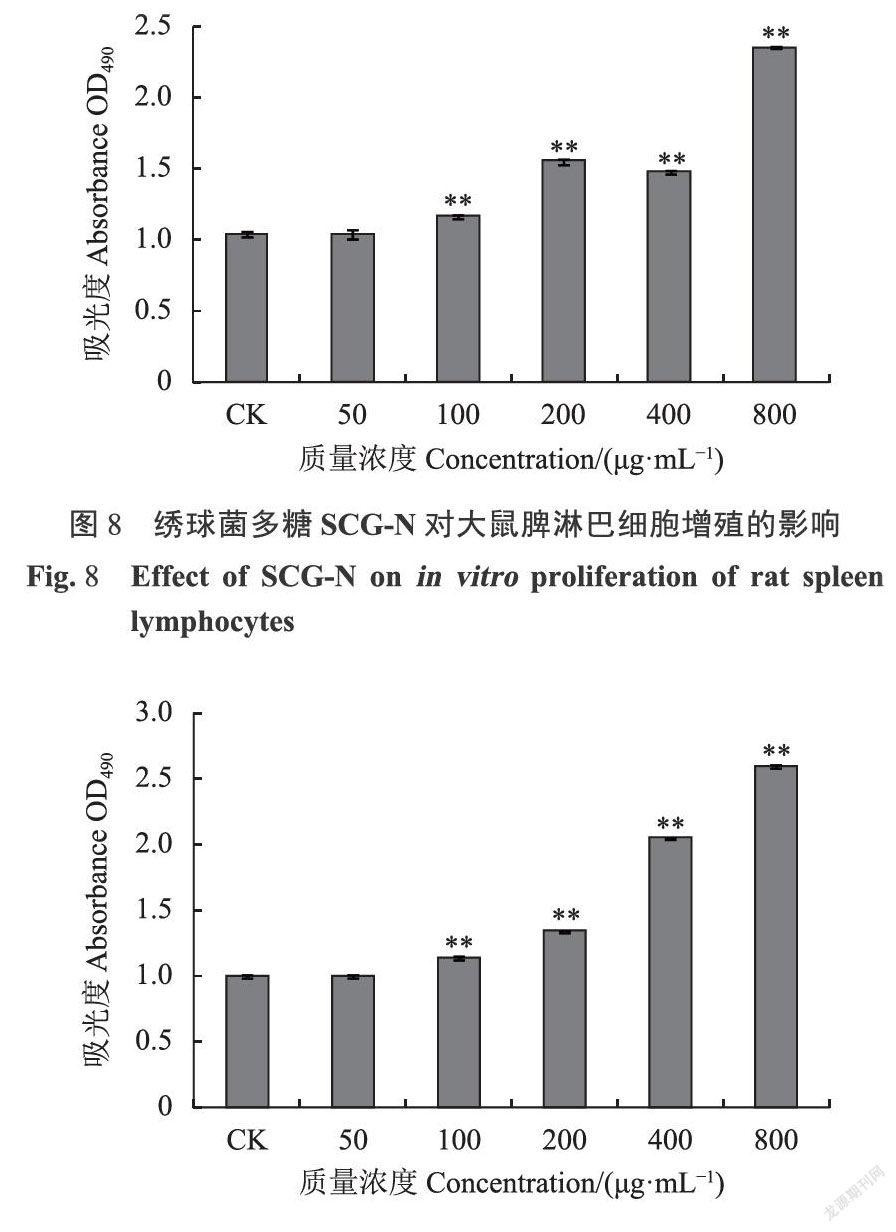
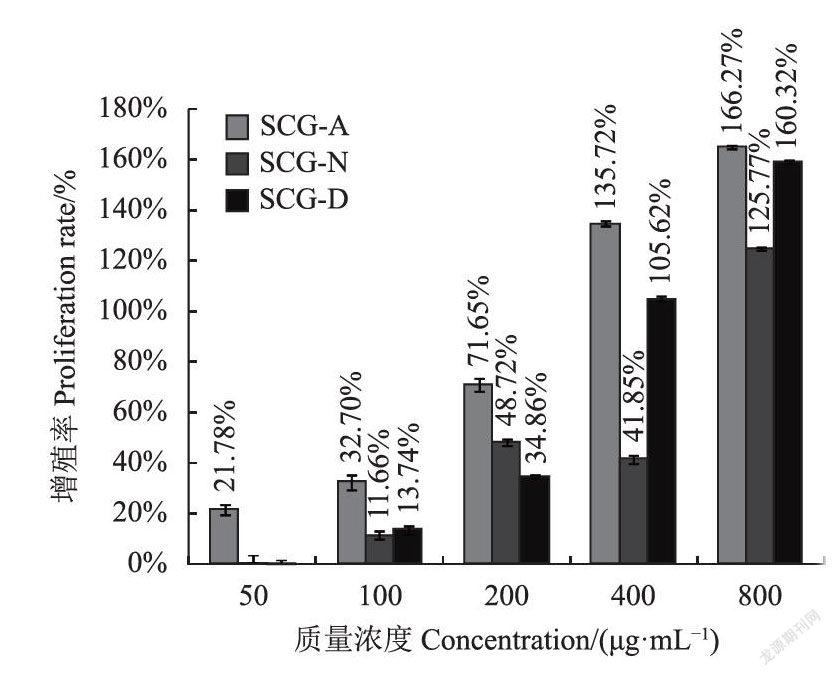
检测结果显示,分子量400KDa左右的SCG-A(图7)、SCG-N(图8)、SCG-D(图9)均表现出不同程度的促进脾淋巴细胞增殖的活性。SCG-A对大鼠脾淋巴细胞的刺激作用较好,在最低质量浓度50ug·mL下仍可以观察到明显的促进作用,增殖率为21.78%(图10),且在50-800ug·mL的范围内存在明显剂量依赖关系,相比之下SCG-D、SCG-N的50ug·mL低质量浓度处理组的刺激效果不显著。
3讨论与结论
在之前的研究中,绣球菌被发现是一种很好制备抗肿瘤葡聚糖的原料,其所述的活性多糖为采用碱溶液提取的难溶性或不溶性多糖,结构分析结果表明这些多糖的一级结构是6-支化的β-1,3-葡聚糖,大约每3个主链单元中有一个分支.本研究从绣球菌冻干品粉末中提取到了一种水溶性多糖,该多糖不同于之前所述的β-1,3-葡聚糖,而是一种α-1,4-葡聚糖,并具有1-6分支结构。同时前期研究表明在烘干品热水提取物中几乎不含有SCG-A,可见SCG-A较容易被生物体降解,这可能与其类似支链淀粉的结构特点相关。
通过大鼠脾淋巴细胞的体外刺激试验对热水提取的绣球菌水溶性多糖及其分离组分的免疫活性进行了检测,检测结果显示冻干品提取的分子量在400kDa左右的多糖表现出了促进大鼠脾淋巴细胞增殖的免疫活性,通过离子交换层析从冻干品多糖中进一步分离得到了SCG-A和SCG-N两个多糖组分,活性检测结果显示二者均有活性,其中酸性多糖SCG-A的活性最强,且剂量依赖关系明显,其含量约占所提得冻干品多糖总量的1/3。
目前为止,我们知道SCG-A的主链结构应为α-1,4-葡聚糖,并含1-6分支结构,但具体的糖链重复单元结构还有待进一步通过甲基化分析和2D-NMR进行确认。同时,其生物学功能也有待进一步的检测和挖掘。
