大气和土壤增温对杉木幼苗叶片功能性状的影响
2019-09-04闫晓俊钟波元陈廷廷张礼宏宋涛涛陈光水
闫晓俊,钟波元,陈廷廷,张礼宏,宋涛涛,陈光水,*
1 福建师范大学地理研究所, 福州 350007 2 福建师范大学湿润亚热带生态地理过程教育部重点实验室, 福州 350007
由于人类活动的加剧,导致全球和区域温度升高,预计在本世纪末全球温度将升高0.3—4.8℃[1]。并且全球气候模型也预测热带亚热带森林区未来将增温3—6℃[2]。温带森林区或苔原区的增温幅度可能大于亚热带森林区,但更多研究发现,全球气候变暖对亚热带森林的生产力、物质循环和生态系统功能产生了重要影响,亚热带森林对全球变暖的响应可能更为敏感脆弱[3]。杉木(Cunninghamialanceolata)为硬叶植物,常绿乔木,属于亚热带树种,喜光喜温喜湿润,忌风忌旱,喜土壤肥沃,不耐严寒和盐碱,叶片为披针形或条状披针形,革质、坚硬等[4]。杉木是生长速度快,经济价值高的用材树种,在世界人工林面积中占重要比例,约6.5%,对我国林业生产和森林碳汇具有重要意义[5]。杉木人工林对增温的响应将为我们科学预测在全球气候变化背景下,杉木人工林通过哪种方式的调整更好地适应变暖环境,以及人工林生产力、碳汇功能作何变化。
叶片的主要功能是帮助植物进行光合作用,是生态系统初级生产者的能量转换器;其功能性状如形态特征、生理代谢特征、化学计量学特征、物候特征等都会对植物的分布与行为产生直接影响,进而影响群落生态系统结构与功能[6]。气候变化对叶片功能性状变异具有重要作用,反映了植物对环境的适应性、自我调节及由此形成的生存、生长、繁殖策略[7]。因此全球变暖对叶片功能性状的影响将直接关系到森林对未来气候变化的适应性。光合作用是植物赖以生存的基础,也是地球碳氮循环的重要过程之一,了解光合作用对模拟增温的响应,可以使我们更好估计在气候变暖背景下,植物自身生理代谢特性、生长及生态系统碳循环的可能变化[8]。叶面积在叶片性状研究中最普遍、使用频率最高,直接影响着植物对光和碳的获取能力。比叶面积同叶片厚度和叶片干物质含量有关,同物种资源获取、潜在生长率和叶片寿命相关性较强,在某种意义上反映了叶片获取光的效率和自我保护能力[9]。而目前关于模拟增温对叶片形态性状和光合作用的研究存在多种结论,有正影响、负影响或没有影响[10- 11]。氮(N)、磷(P)是限制植物生长的最主要的两种元素,叶片N、P浓度的高低不但可以反映叶片的生理功能,同时也是评估植物营养状况的重要指标。有研究表明,在全球尺度上叶片N、P浓度与年均温表现为负相关关系[12]。而在区域尺度或小尺度上,模拟增温对叶片N浓度的影响结果呈现多样化,有增加[13- 14],降低[15- 17],或无显著影响[18- 19]。热带亚热带土壤发育高度风化,养分极易被淋溶且P易和铁铝氧化物结合,植物对P元素的吸收有限,而植物叶片磷浓度变化对增温响应研究还相对较少。
目前,关于植物对增温的响应研究主要分布在中高纬度地区[20],而在我国的亚热带森林的增温试验开展较少。而且目前模拟增温试验采用不同的增温方式,如红外增温、土壤电缆增温、开顶箱(OTC)被动式增温等,这些不同的增温方式对植物地上和地下部分的增温效果不一,即要么主要对地上部分增温(如红外增温和OTC增温),要么主要对地下部分增温(如土壤电缆增温),不同的增温方式可能造成了研究结果的差异;而考虑对整个植物系统地上和地下同时增温的研究很少[21],而只有地上和地下同时增温才能更好地模拟未来变暖的环境。因此在亚热带开展增温实验,特别是同时大气和土壤增温对杉木幼苗叶片功能性状的影响,揭示其对全球气候变暖的响应与适应机制,为预测亚热带人工林森林生产力和碳汇潜力提供基础数据。根据前人所作研究,本论文做出如下假设:(1)土壤增温和大气增温可能会促进杉木幼苗叶片净光合速率和水分利用效率的增加;(2)大气增温可能增加杉木幼苗叶片含水率和比叶面积,减小叶片厚度,但土壤增温对此并无影响;(3)土壤增温和大气增温可能会增加叶片氮浓度。
1 材料与方法
1.1 试验地概况
实验地点坐落在福建三明森林生态系统与全球变化研究站陈大观测点(26°19′ N,117°36′ E)。本区域气候属于中亚热带季风气候,年均温19.1℃,年降雨量1749 mm,每年3—8月多发生降雨,年均蒸发量较大,为1585 mm,相对湿度81%。红壤和黄壤为地带性土壤,杉木经济林种植广泛,区内物种多样性丰富。
1.2 试验设计
试验处理设置为大气增温(AW,两水平:+0,+OTC增温)×土壤增温(SW,两水平:+0,+4℃)双因子实验,包括对照处理(CT),土壤增温处理(W),大气增温处理(OTC)和大气和土壤同时增温处理(OTC-W),总共4个处理,且每个处理12个重复。每个根箱种植1株高度和地径相似的杉木幼苗,共计 48个根箱,根箱长、宽、高为50、50、60 cm,四周被钢化玻璃覆盖,为了避免玻璃与土壤表层所生根系受到外界环境条件的影响,在根箱四面安装隔热板。根箱底部未封闭且置于地面土壤上,这有利于根系向箱下土壤拓展延伸,以免受生长空间约束。根箱内土壤取自附近杉木林原位土,取土时分为3层,分别是0—20 cm、20—40 cm和40—60 cm,在挑拣杂物后将土壤尽可能混匀,达到消除土壤异质性的目的,接着分层填回根箱,并通过压实法调整土壤容重使尽可能与取土杉木林土壤条件保持一致[22]。
土壤增温方案是在每个根箱平行安装布设电缆,为确保增温的均匀性,电缆处于土壤10 cm处,间隔20 cm,并在最外围环绕一圈电缆。同时在每个根箱放置5个温度传感器(德国JUMO)和2个水分传感器(美国Decagon),其中将3个温度传感器埋在10 cm处,另外2个放在表层5 cm和深层20 cm处;水分传感器置于土层10 cm处以便测定含水率[22]。温度和水分传感器均布设在两条电缆线中间位置。土壤增温控制是通过采集无土壤增温处理(CT、OTC)根箱和有土壤增温处理(W、OTC-W)根箱的土壤温度进行对比,具体土壤增温技术环节详见参考文献[23]。大气增温方案是搭建2个开顶箱(Open-top chamber,OTC)进行被动式增温,高度达到4.8 m。在OTC内不同高度处安装温湿度探头,利用温湿度探头对OTC内外温度和湿度进行连续自动观测。2015年11月份开始大气增温(OTC建成),2016年3月开始土壤增温。各处理的小气候指标和树高生长量见图1。
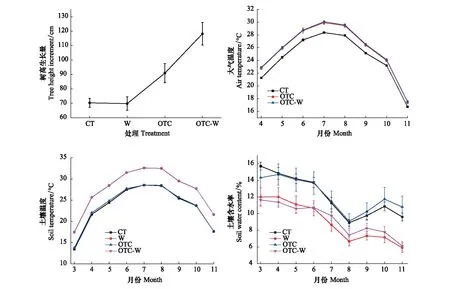
图1 不同处理的气候指标及树高生长量Fig.1 Climatic indexes and tree height growth of different treatmentsCT:对照处理 Contrast treatment; W:土壤增温处理 Soil warming treatment;OTC:大气增温处理 Air warming treatment; OTC-W:土壤和大气同时增温处理 Both soil warming and air warming treatment
1.3 样品采集与指标测定
光合作用在2016年4、7、11月采用CIRAS- 3便携式光合仪测定,每次选择不同处理苗木3株。在每棵幼苗中上部向阳方位的一年生枝条上选择3片健康的一年生叶,利用测定时外部环境具有的温湿度值和二氧化碳浓度数值作为参数,并将光强设定为饱和光强(1500 μmol m-2s-1)测定叶片净光合速率(Pn)及水分利用效率(WUE)等指标,时间集中在9:00—11:00。测定过程中,在仪器稳定1—2 min后进行多点读取数据。光合作用测定完毕后把叶片摘下,利用数字化扫描仪Espon scanner 分析叶面积,然后进行数据的换算。
在增温后近1年的2016年11月,在每棵幼苗中上部四周(包括东西南北)随机选择8—10枚健康、成熟的当年生叶。然后对叶片进行形态特征及养分含量的测定。
叶厚度(Leaf thickness, LT,mm),利用游标卡尺在幼苗叶片上部、中部、下部依次进行测量读数,然后将3个数值取平均数作为叶片实际厚度。
叶面积(Leaf area, LA,cm2),通过数字化扫描仪Espon scanner 得到扫描图片,然后采用Win-RHIZO(Pro 2009b)分析系统对扫描图进行分析处理,从而得到叶面积。
比叶面积(Specific leaf area, SLA, cm2/g),即单位干重的叶面积,SLA=LA/叶干重。
叶组织密度(Leaf tissue density, LTD, g/cm3),即单位体积的叶片干重,LTD=叶干重/叶体积。
叶含水率(Leaf water content, LWC, %),用天平称量采集的杉木幼苗叶片作为鲜重,杀青后,将样品放入65℃烘箱烘至恒重后,用天平称取作为干重,LWC=[ (鲜重-干重)/干重×100]。
叶碳(C)、氮(N)、磷(P)浓度,利用球磨仪将叶片样品进行粉碎直至碎末,采用Elementar Vario EL Ⅲ元素分析仪测定叶片C浓度、N浓度;经过硫酸-高氯酸消煮后,使用Skalar San++连续流动分析仪上机测定叶片P浓度。
1.4 数据处理
采用双因素方差分析法(two-way ANOVA)检验大气增温、土壤增温及二者交互作用对各测定时间净光合速率、水分利用效率,以及叶片形态和化学计量学特征的影响;当存在交互作用时,进一步采用LSD多重比较分析不同处理间的差异。利用Excel 2003和SPSS 19.0软件对数据进行统计分析,将P=0.05作为显著性水平标准。利用Origin 9.0软件制作相关图表。
2 结果与分析
2.1 叶片光合特征
双因素方差分析表明,4月和7月大气增温、土壤增温及其大气增温和土壤增温交互作用对杉木幼苗1年生叶片的净光合速率和水分利用效率均没有显著影响(P>0.05),但土壤增温对11月的叶片净光合作用速率有显著影响(P<0.05),大气增温和土壤增温对11月的水分利用效率均有显著影响(P<0.05)。通过对11月不同处理进行LSD多重比较发现, OTC-W处理叶片净光合速率显著高于CT;且OTC-W处理的水分利用效率显著高于CT和W处理(图2)。
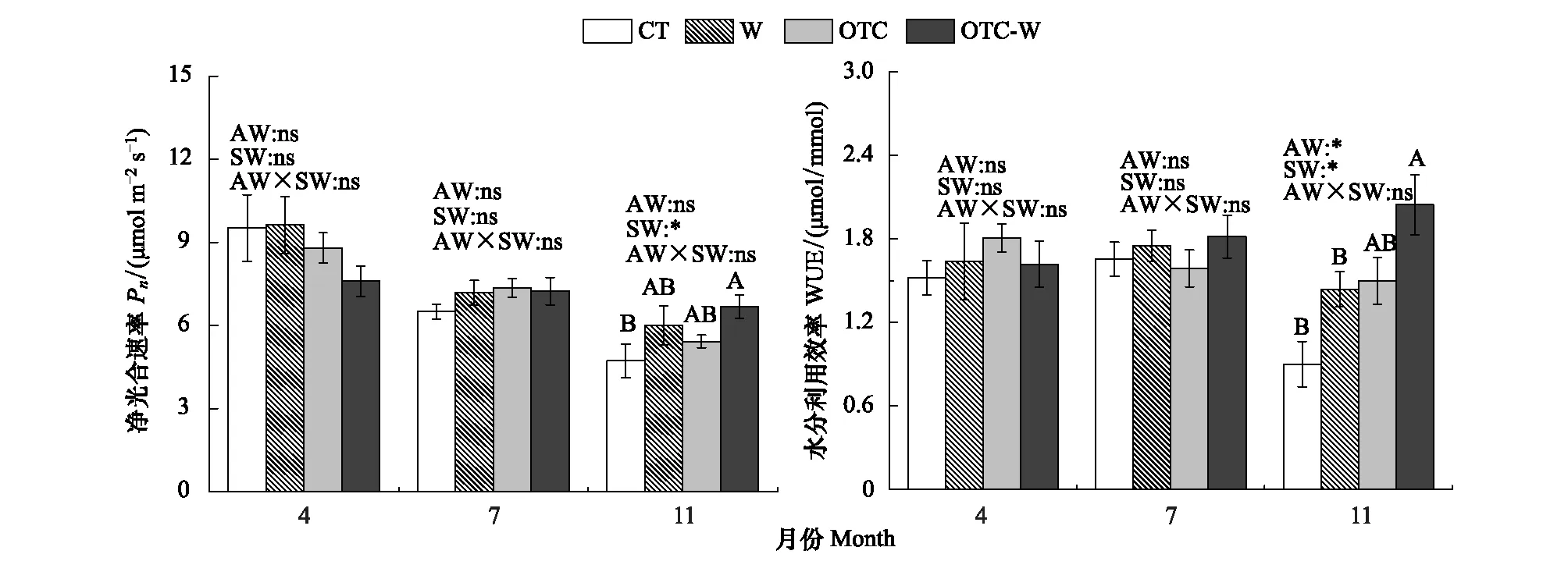
图2 不同增温方式对杉木幼苗净光合速率及水分利用效率的影响Fig.2 Effects of warming on photosynthetic rate and water-use efficiency of Cunninghamia lanceolata seedlingsAW:大气增温 Air warming; SW:土壤增温 Soil warming; AW×SW:土壤与大气同时增温 Both soil warming and air warming;*: P<0.05 ns:无显著影响 not significent; 不同大写字母表示相同季节不同处理差异显著
2.2 叶片形态特征
根据图3可知,通过双因素方差分析发现,大气增温对杉木幼苗叶片的比叶面积、叶含水率起到了显著增加作用(P<0.05),且大气增温显著降低了杉木幼苗叶片的厚度(P<0.05),大气增温对叶面积、组织密度没有产生显著作用(P>0.05),土壤增温及土壤和大气同时增温对杉木幼苗叶片的各个形态性状都没产生显著作用(P>0.05)。该结果亦表明大气增温对比叶面积的影响,主要通过对叶厚度的调整所致(图3)。
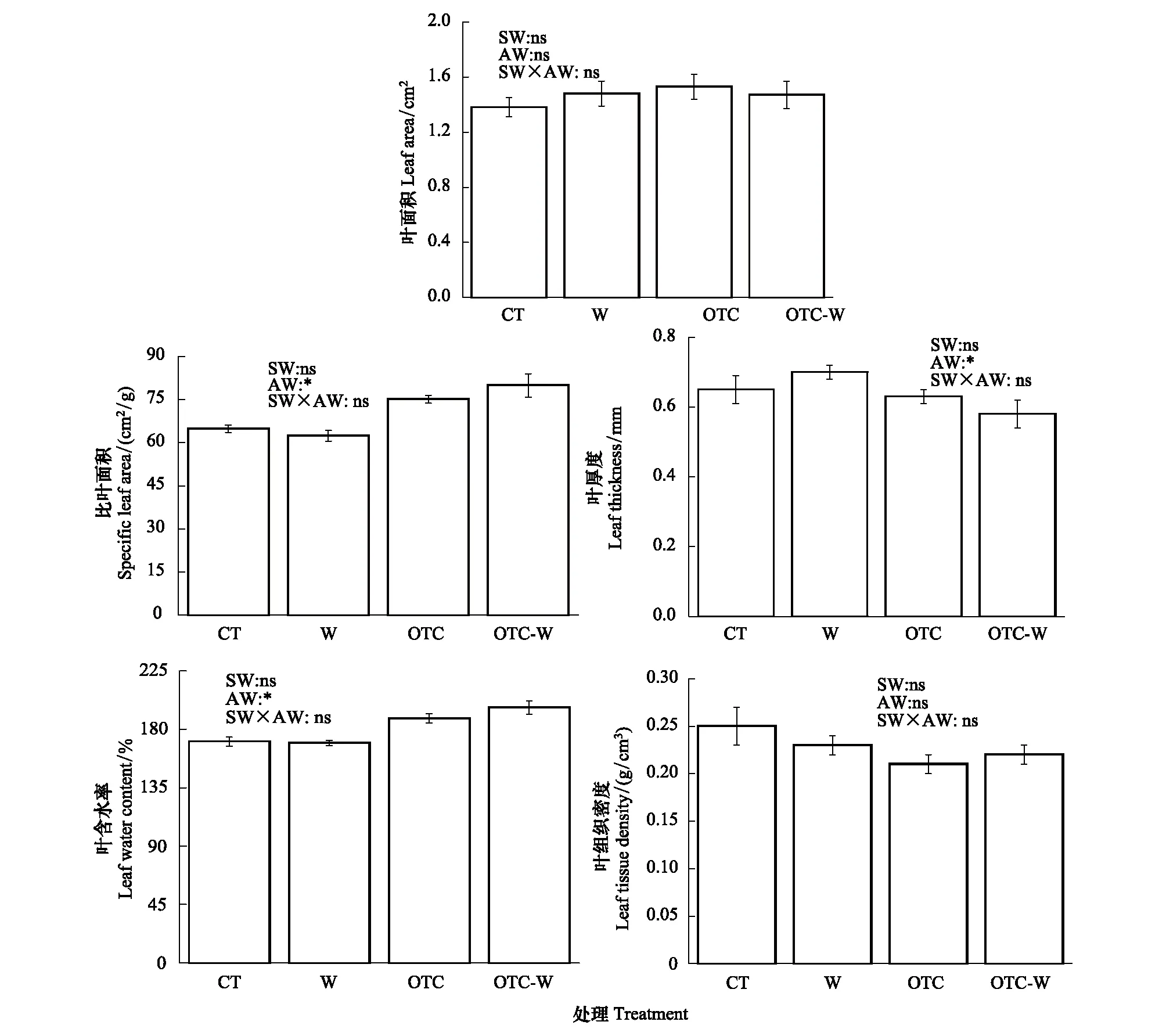
图3 不同增温方式对杉木幼苗叶片形态结构特征的影响Fig.3 Effects of warming on leaf morphological traits of Cunninghamia lanceolata seedlings
2.3 叶片元素化学计量学特征
从图4可以看出,双因素方差分析结果表明,土壤增温对杉木幼苗叶片的碳浓度、氮浓度、磷浓度及碳氮比、碳磷比都产生显著作用 (P<0.05),但对氮磷比没产生显著作用 (P>0.05);大气增温对叶片氮浓度、碳氮比、氮磷比具有显著作用(P<0.05),对叶片碳浓度、磷浓度、碳磷比没有产生显著影响(P>0.05);土壤增温与大气增温的交互作用只对叶片磷浓度产生显著作用(P<0.05),对其他指标无显著影响(P>0.05)。进一步观察可知,土壤增温显著增加了杉木幼苗叶片的碳浓度、氮浓度,显著降低了碳氮比和碳磷比;大气增温显著增加了氮浓度和氮磷比,显著降低了叶片的碳氮比。通过对叶磷浓度进行LSD多重比较发现,OTC-W处理的叶磷浓度显著高于CT和其他处理(图4)。
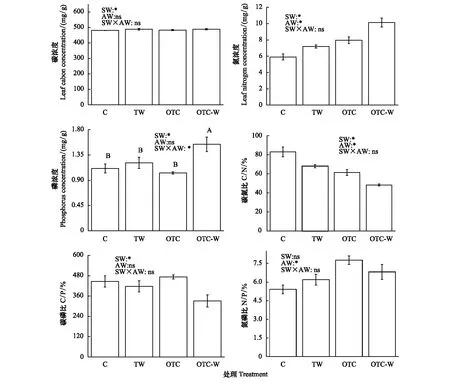
图4 不同增温方式对杉木幼苗叶片化学计量学特征的影响Fig.4 Effects of different warming ways on leaf stoichiometry of Cunninghamia lanceolata seedlings不同大写字母表示不同处理之间差异显著Different capital letters meant significant difference among different treatments
3 讨论
3.1 大气和土壤增温对叶片光合特征的影响
光合作用是植物赖以生存的基础,也是地球碳氮循环的重要过程之一,与温度紧密相关;有研究发现,当温度低于光合作用最适温度时,光合速率与温度表现为正相关,当温度高于最适温度后,光合速率则随着温度的升高而降低,不利于光合作用[24]。本研究中大气增温对叶片净光合作用速率没有显著影响,而土壤增温仅在11月对叶片净光合速率有显著影响。这与假设(1)大气增温和土壤增温均可能促进净光合速率的升高有出入。一些研究也得到了类似结论,如Yang等[17]研究发现,大气增温对青藏高原小蒿草单位面积的净光合速率无显著影响;Li等[10]对亚热带树种的空间置换增温研究发现,红枝蒲桃的光合速率没有发生显著变化。此外,也有一些研究得出与本研究不同的结果。如Niu等[8]采用红外辐射加热模拟实验研究则发现增温显著降低克氏针茅净光合速率,可能是因为在水分胁迫情况下,增温对光合作用的影响减弱。徐振锋等[25]通过研究发现,大气增温促使糙皮桦幼苗净光合速率增加;其研究发现增温显著增加了最大羧化效率和电子传递率,从而促进了净光合速率。本研究中4月和7月大气增温、土壤增温及其交互作用对叶片净光合作用速率没有显著影响,可能是4月和7月气温本身已处于叶片进行光合作用的适宜温度,因而气温升高对光合作用的影响不显著;同时4月和7月的土壤温度本身也处于土壤微生物活动的适宜范围,因而土壤增温对于根系的养分、水分吸收的影响较小,进而对光合作用的间接作用也小。11月时虽然大气温度较低,但此时OTC的大气增温效应较小(图1),因而大气增温对叶片净光合速率没有显著影响;而此时土壤温度也较低,土壤增温可能明显促进了根系的活动以及水分、养分的吸收,从而可能间接地促进了光合作用。
了解植物的水分利用效率(WUE),能够更好地认识植物内在的耗水原理及使用效率,与生态系统碳循环和水循环有关,一般而言,植物水分利用效率越高,耐旱性越好。植物蒸腾过程受到了叶片气孔导度的影响,温度升高对叶片净光合速率和蒸腾速率的增幅有差异,因此水分利用有效率对温度的响应存在正、负之分[26]。本研究中土壤增温和大气增温均增加11月杉木幼苗叶片的水分利用效率。此结果印证了土壤增温和大气增温会促进WUE这一假设。这与Niu等[27]研究发现增温对克氏针茅叶片WUE没有产生显著作用的结果明显不同。可能因为当地此段时间降水较少,土壤增温导致土壤含水率严重降低;杉木幼苗为避免受到水分胁迫,从而提高水分的利用效率来维持自身生长。
3.2 大气和土壤增温对叶片形态特征的影响
本研究中大气增温显著增加杉木幼苗比叶面积(SLA)和含水率(LWC),显著降低了叶片厚度,但并没有影响叶面积。这与本研究提出的假设(2)相吻合。一些研究得出类似结论,如Yang等[17]的研究发现,大气增温显著降低小蒿草的单位面积叶干重,表明其SLA显著增加;杨兵等[28]通过控制环境生长室模拟增温的研究发现,增温显著增加岷江冷杉SLA。本研究中大气增温对SLA的促进作用,可能是叶片厚度降低所致,因为叶片的组织密度并没有发生显著变化。此外,Hudson等[18]研究表明,大气增温显著增加冻原植物Cassiopetetragona叶面积,但SLA却显著降低,可能原因是叶面积增大导致组织构建成本升高;羊留冬等[29]研究也发现,利用红外线加热器增温虽然增大了贡嘎山峨眉冷杉的叶面积,但显著降低SLA。而Suzuki和Kudo[30]研究表明,OTC模拟增温没有显著影响植物叶片大小和SLA,这可能与不同植物自身对增温的响应模式有关。有研究表明SLA随着LWC的升高而增加[31],本研究肯定了这一论断,在大气增温条件下杉木幼苗SLA和LWC都呈增加趋势。SLA和LWC的增加表明了大气增温下杉木幼苗叶片生理功能的提升。而土壤增温对二者皆无显著影响,可能原因是土壤增温没有显著改变土壤环境因子,导致叶片的表型可塑性不明显[32],也可能是土壤增温直接影响杉木幼苗地下根系生长,而对幼苗地上器官(叶片)影响很小,侧面反映出不同增温方式对杉木叶片形态特征影响不一。
3.3 大气和土壤增温对叶片元素化学计量学特征
此次研究发现土壤增温和大气增温都对杉木幼苗叶片氮浓度(LNC)表现为显著增加作用,同时而对碳氮比(C∶N)则表现为显著降低;此外,大气增温对叶片氮磷比(N∶P)产生显著增加作用。这印证了假设(3)。一些研究得出相似结论,如Butler等[13]通过长达7年对红枫树的土壤增温研究发现,在增温2年后,叶片N浓度显著升高,可能是土壤氮矿化的增加导致叶片氮含量增加。 Rodgers等[14]研究也发现不同梯度的增温显著增加了LNC和显著降低叶片C∶N,表明在增温背景下植物对氮养分的获取吸收明显增多,且在分配过程中将更多的氮元素运输到叶片。可能是土壤增温使土壤有效氮含量提高,并且大气增温影响叶片的气孔导度,使得蒸腾作用加快,促进了植物对土壤N元素的吸收。两种增温方式下杉木幼苗叶片C∶N显著降低,可能原因是LNC显著升高,而碳浓度基本保持不变。但是Reich和Oleksyn[12]发现在全球尺度上,叶片LNC随着温度上升反而下降。同时野外模拟增温研究亦表明温度与叶片LNC表现为负相关[15,17,33]。一些研究发现增温促进植物生长量的提高,而植物对氮元素的吸收速率与植物生物量的增加不相匹配,从而叶片氮浓度增加不显著[15],也或许是增温促进植物的氮利用效率的提高,进而使植物LNC下降。另外增温使土壤水分减少,更加干旱,植物在受到水分胁迫的情况下,作出减少吸收氮元素的反映[16- 17]。也有研究表明增温对植物叶片氮浓度影响不显著[18- 19],这有可能是在土壤有效氮含量一定的情况下,为了生存生长,土壤微生物与植物间存在激烈的竞争关系,植物组织可能没有及时充分地获取氮养分[34];亦或是增温影响了氮资源地上、地下的分配,因为增温后细根活性加强,导致细根对氮元素的吸收能力增强,使得氮分配到叶片的含量较少[35]。本研究中,只有土壤和大气同时增温处理对杉木叶片P浓度才有显著增加作用。可能是因为两者同时增温促进了杉木幼苗的生长,这样苗木可以分配更多的光合产物以促进地下根系的生长、提高菌根菌侵染率或增加根系分泌物的分泌,从而提高P的吸收以供地上部分生长。而江肖洁等[36]在长白山开顶箱模拟增温发现,增温使长白山苔原植物生长受P元素的限制更加明显。Yang等[17]通过不同水平模拟增温试验发现,大气增温对植物叶片的P浓度显著降低,增温幅度更大的处理(+4.4 ℃)根系P浓度却表现为显著升高,表明增温使P元素在地上地下的分配过程具有差异性。
N∶P通常被用来判定植物受何种养分限制,有研究表明当植物N∶P<14时,植物生长繁殖受到氮元素制约;N∶P>16时,说明植被生长受磷元素的约束;14 土壤增温促进了11月的杉木叶片净光合作用速率和水分利用效率;而大气增温促进11月叶片的水分利用效率,显著增加比叶面积和叶含水率,并降低叶厚度。土壤增温、大气增温均显著增加杉木叶氮浓度,但只有同时大气增温和土壤增温才显著提高叶磷浓度。4 结论
