洞庭湖不同水位梯度川三蕊柳和短尖薹草土壤有机碳垂直分布特征
2019-08-29边华林杨广军牛艳东邓嘉俊李有志丁小慧周巧巧
边华林,杨广军,牛艳东,邓嘉俊,李有志,丁小慧,姚 欣,周巧巧
(1.湖南农业大学生物科学技术学院,湖南 长沙 410128;2.岳阳市林业科学研究所,湖南 岳阳 414000;3.湖南省林业科学院,湖南 长沙 410004)
湿地是陆地和水域之间的过渡地带,碳储量占全球陆地生态系统的12%~20%[1]。土壤是湿地碳库的主体,占据碳储量的95%,对湿地碳循环具有重要作用[2]。植被通过光合作用固定二氧化碳是土壤碳的主要来源,其固碳量与植被类型密切相关[3-4]。草本和木本是植物的2种主要生活型,其生态功能差异明显[5]。研究发现,草本植物灰化薹草(Carexcinerascens)年固碳量可达1 140 g·m-2[6],远高于木本植物红树林(444.27 g·m-2)[7]。在黄河三角洲,盐地碱蓬(Suaedasalsa)等草本植物固碳量也高于柽柳(Tamarixchinensis)、白刺(Nitrariatangutorum)、杞柳(Salixintegra)等木本植物[8]。此外,草本植物和木本植物固碳后形成的碳水化合物存在差异(如纤维素、半纤维素、木质素等),可能也会导致植物凋落物或残体分解速率不同,从而影响土壤碳输出[9]。在东北草本修氏薹草(Carexschmidtii)湿地,土壤以富里酸为主,占总腐殖质的46.23%,而在落叶松(Larixgmelinii)湿地,土壤以胡敏酸为主,占总腐殖质的49.06%[10]。研究发现,木质素含量低的草本植物花拳参(Gutierreziasarothrae)、格兰马草(Boutelouagracilis)分解速率显著高于木质素含量高的木本植物西黄松(Pinusponderosa)、食松(Pinusedulis)、北美樱桃核桧(Juniperusmonosperma)[11]。可见,木本和草本植物的类型差异势必影响土壤碳库。
在湿地生态系统中,洪水携带的大量泥沙也是土壤碳库不可忽视的重要来源之一[12]。随着湿地高程的降低,水流量增大,泥沙淤积量也随之增加。在三峡库区,2006—2015年泥沙淤积量在170 m高程区为650.00 kg·m-2,162 m高程区增加到938.96 kg·m-2[13]。木本和草本植物因形态特征的差异影响水流速度,从而影响泥沙淤泥。监测发现,与芦苇(Phragmitesaustralis)群落相比,杨树(Populus)林水流速度增大,泥沙量增加[14]。在洪水季节,木本植物川三蕊柳(Salixtriandroides)的水流速率小于草本植物短尖薹草(Carexbrevicuspis),因此前者泥沙沉积速率较高,这可能导致两者之间土壤碳含量的差异[5]。此外,随着湿地高程的降低,土壤含水量逐渐增加,进而影响土壤碳的分解。研究发现,在常年积水的芦苇湿地,土壤胡敏酸和富里酸碳含量随着土壤含水量的升高而增加,而在季节性积水的芦苇湿地,土壤胡敏酸和胡敏素碳含量则与土壤含水量呈显著正相关关系[10]。因此,湿地植物对土壤碳库的影响可能与高程等生境因子有关,需要开展系统的对比研究。洞庭湖为我国第二大淡水湖泊,分布有短尖薹草、辣蓼(Polygonumhydropiper)、芦苇(Phragmitesaustralis)等草本植物,同时也保留川三蕊柳等木本植物。然而,随着洞庭湖湿地环境的变化,川三蕊柳退化为薹草群落。因此,研究选取洞庭湖不同高程区域的川三蕊柳和短尖薹草为研究对象,对比土壤有机碳及腐殖质碳组成,比较木本和草本植物群落土壤碳库的区别,评估川三蕊柳群落退化对洞庭湖土壤碳库的影响,为洞庭湖川三蕊柳保护和湿地资源管理提供理论参考。
1 研究区域
1.1 研究区域概况
洞庭湖位于荆江南岸,面积为2 625 km2(28°44′~29°35′ N,111°53′~113°05′ E),高程28~35 m(图1)。该区域属于典型的亚热带季风气候区,年平均气温16.5~17 °C,1月平均气温最低,为4.2 ℃,7月平均气温最高,为29 °C。年平均降水量为 1 250~1 450 mm,蒸发量为1 270 mm。5月开始进入主汛期,10月结束,平均水位落差12~14 m。

图1 洞庭湖湿地采样点位置示意Fig.1 Location of investigated plots in the Dongting Lake wetlands
1.2 研究对象
川三蕊柳,属杨柳科柳属小乔木,高2~5 m,为洞庭湖湿地原生木本植物,因其分枝多、枝条柔软,具有消浪护堤、生物多样性保育等重要生态功能[15-16]。短尖薹草属莎草科薹草属草本植物,高0.2~0.6 m,为洞庭湖湿地建群种优势植物之一,是候鸟、鱼类等野生动物的重要食物来源。
2 研究方法与数据处理
2.1 研究方法
2017年12月(洪水退后),在东洞庭湖区洲滩地(29°21′50″ N,112°54′53″ E)选取3种水位梯度的川三蕊柳群落,以邻近10 m左右的草本植物短尖薹草为对照。在每种植物群落中央选取3个20 m×30 m的调查样方。在每个调查样方内挖取3个口径为50 cm的柱形洞,实测地下水位深度,3个水位梯度植物群落平均地下水深分别为-60 cm(低水位,水位在土壤表层60 cm以下)、-40 cm(中水位,水位在土壤表层40 cm以下)和-20 cm(高水位,水位在土壤表层20 cm以下)。同时,分层采集(0~20、>20~40、>40~60、>60~80 cm)土壤样品,将同一水位梯度同一植物群落样品混合,自封袋装好带回实验室。在2018年12月,在每个川三蕊柳群落内选取3个20 m×30 m的调查样方,统计样方内川三蕊柳数量,选取标准木3株伐倒,测定其树叶、树枝、树干和树根生物量,并在每个样方内选取3个1 m×1 m的草本样方,测定地上与地下部分生物量;在每个短尖薹草群落内选取3个1 m×1 m的调查样方,测定地上与地下部分生物量。
将从野外带回的土壤样品小心剔除植物残渣,自然晾干,过120 mm孔径筛后,分析理化性质。土壤有机碳含量采用重铬酸钾氧化外加热法测定,土壤腐殖质各组分(胡敏酸、富里酸、胡敏素)含量采用焦磷酸钠-氢氧化钠溶液提取法测定[17]。
2.2 数据处理
利用SPSS 21.0数据分析软件对不同植被类型、水位深度、土壤深度下土壤有机碳、胡敏酸碳、富里酸碳和胡敏素碳含量进行方差分析,显著性水平设为0.05。对同一土层深度下,3种水位和2种植物群落之间的土壤有机碳及各组分含量进行Tukeys-b(K)式多重比较,显著性水平设为0.05。在分析前对数据进行方差齐性检测,对于不符合齐性检测的数据进行对数转换。
3 结果与分析
3.1 植物群落生物量
就同一植物群落而言,生物量以中低水位区的较高,高水位区的较低。如川三蕊柳群落,低水位区和中水位区的生物量分别是高水位区的3.7和3.3倍。就同一水位梯度而言,川三蕊柳植物群落生物量总体上高于短尖薹草(表1)。

表1 3种水位梯度下川三蕊柳和短尖薹草生物量Table 1 Plant biomass of Salix triandroides and Carex brevicuspis under three ground water level gradients kg·m-2
数据为平均值±标准差。同一列数据后英文小写字母不同表示不同水位梯度间群落生物量差异达显著水平(P<0.05)。
3.2 土壤有机碳
由表2~3可知,在同一植物群落中,3种水位梯度下土壤有机碳含量差异显著(F=112.378,P<0.001)。川三蕊柳群落表层0~20 cm土壤有机碳含量以高水位区较高,其余3层土壤有机碳含量以中水位和低水位区较高。短尖薹草群落土壤有机碳含量除>20~40 cm土壤外,其余3层均以中水位区较高。
在同一水位梯度下,2种植物群落之间土壤有机碳含量差异显著(F=20.780,P<0.001)。在低水位区,4种土壤深度下有机碳含量均以川三蕊柳群落较高。在中水位区,除>40~60 cm土壤2种植物群落之间土壤有机碳含量无显著差异外,其余3层土壤有机碳含量均以短尖薹草群落较高。在高水位区,0~20 cm土壤有机碳含量以川三蕊柳群落较高,>20~40和>40~60 cm土壤以短尖薹草群落较高,而2种植物群落之间>60~80 cm土壤有机碳含量无显著差异。方差分析结果表明,土壤深度对有机碳含量有显著影响(F=4 267.562,P<0.001)。
3.3 胡敏酸碳含量
由表2~3可知,在川三蕊柳群落中,0~20 cm土壤胡敏酸碳含量以高水位区较高,>60~80 cm土壤以高水位区较低,而>20~40和>40~60 cm土壤3种水位下胡敏酸碳含量无显著差异。短尖薹草群落表层0~20和>20~40 cm土壤胡敏酸碳含量以中水位区较高,>60~80 cm土壤以高水位区较低,>40~60 cm土壤3种水位下胡敏酸碳含量无显著差异。方差分析结果表明,水位深度对土壤胡敏素碳含量无显著影响(F=1.300,P>0.05)。
对低水位区而言,>20~40 cm土壤胡敏酸碳含量以短尖薹草群落较高,其余3层土壤胡敏酸碳含量在2个群落之间无显著差异。对中水位区而言,除>60~80 cm土壤胡敏酸碳含量在2个群落之间无显著差异外,其余3层土壤胡敏酸碳含量均以短尖薹草群落较高。对高水位区而言,0~20 cm土壤胡敏酸碳含量以川三蕊柳群落较高,而>40~60 cm土壤以短尖薹草群落较高,>20~40和>60~80 cm土层胡敏酸碳含量在2个群落之间无显著差异。方差分析结果表明,植被类型对土壤胡敏素碳含量无显著影响(F=0.405,P>0.05),而土壤深度对胡敏素碳含量存在显著影响(F=279.005,P<0.001)。
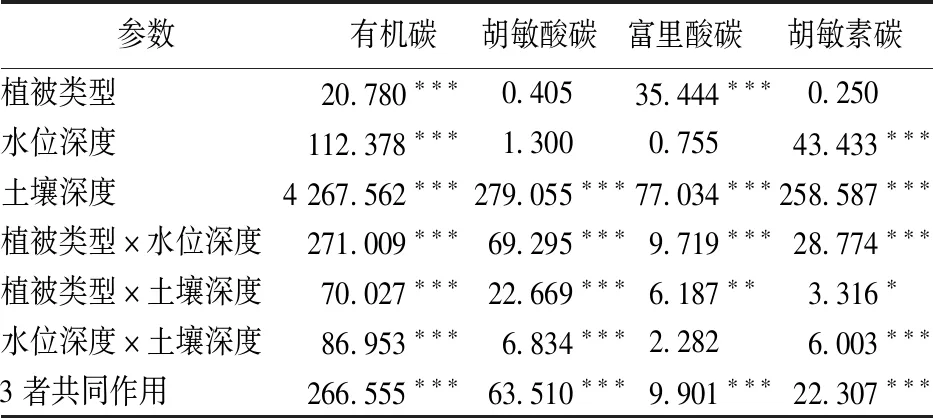
表2 植被类型、水位深度和土壤深度对土壤有机碳和腐殖质碳含量影响的多因素方差分析结果(F 值)Table 2 Repeated-measure ANOVA of soil organic carbon,soil humic acid carbon,soil fulvic acid carbon,and soil humin carbonof Salix triandroides and Carex brevicuspis under three ground water level (F values)
*表示P<0.05;**表示P<0.01;***表示P<0.001。

表3 3种水位梯度下川三蕊柳和短尖薹草土壤有机碳和腐殖质碳含量Table 3 The soil organic carbon,soil humic acid carbon,soil fulvic acid carbon,and soil humin carbon of Salix triandroides and Carex brevicuspis under three ground water level gradients
数据为平均值±标准差。同一土层深度同一指标数据后英文小写字母不同表示某指标差异达显著水平(P<0.05)。
3.4 富里酸碳含量
在川三蕊柳群落中,除>60~80 cm土壤富里酸碳含量表现为中水位区低于高水位区外,其余3层土壤富里酸碳含量在3种水位之间无显著差异。在短尖薹草群落中,除>40~60 cm土壤富里酸碳含量以中水位区较高外,其余3层土壤富里酸碳含量在3种水位之间也无显著差异。方差分析结果表明,水位深度对土壤富里酸碳含量无显著影响(F=0.755,P>0.05,表2)。
在低水位区域,除0~20 cm土壤富里酸碳含量在2种群落之间无显著差异外,>20~40、>40~60、>60~80 cm深度3种水位下土壤富里酸碳含量以川三蕊柳较高。在中水位区,2种植物群落之间土壤富里酸碳含量均无显著差异。在高水位区,0~20和>40~60 cm土壤富里酸碳含量以川三蕊柳较高,而>20~40和>60~80 cm土壤富里酸碳含量在2种植物群落之间无显著差异。方差分析结果表明,土壤深度(F=77.034,P<0.001)和植被类型(F=35.444,P<0.001)均对土壤富里酸碳含量存在显著影响。
3.5 胡敏素碳含量
同一植物群落中,3种水位梯度下土壤胡敏素碳含量差异显著(F=43.433,P<0.001)。川三蕊柳群落0~20 cm土壤胡敏素碳含量以高水位区较高,而其余3层土壤胡敏素碳含量均以高水位区较低。总体上,短尖薹草群落土壤胡敏素碳含量均以中水位区较高。
在低水位区,除>40~60 cm土壤胡敏素碳含量以川三蕊柳群落较高外,其余3层土壤胡敏素碳含量在2个群落之间无显著差异。在中水位区,0~20和>60~80 cm土壤胡敏素碳含量以短尖薹草群落较高,而>20~40和>40~60 cm土壤胡敏素碳含量在2个群落之间无显著差异。在高水位区,0~20 cm土壤胡敏素碳含量以川三蕊柳群落较高,>20~40和>40~60 cm土壤胡敏素碳含量以短尖薹草群落较高,而>60~80 cm土壤胡敏素碳含量在2个群落之间也无显著差异。方差分析结果表明,植被类型对土壤胡敏素碳含量影响不显著(F=0.250,P>0.05),而土壤深度对土壤胡敏素碳含量存在显著影响(F=258.587,P<0.001)。
4 讨论
研究表明,洞庭湖湿地川三蕊柳和短尖薹草2种植物群落土壤有机碳和腐殖质碳(胡敏酸碳、富里酸碳、胡敏酸碳)含量均以0~20 cm土壤较高。张文菊等[18]对洞庭湖草滩地的研究发现,土壤有机碳含量以表层(0~10 cm)最高,之后随着深度增加而降低。在三江平原沼泽湿地中,土壤有机碳含量最大值出现在0~15 cm土层[19-20]。一方面,枯枝、落叶等植物凋落物主要归还于土壤表层,因此该层土壤有机碳含量较高[21];另一方面,有机酸和糖类等根系分泌物是土壤碳的重要来源[22],湿地植物根系分布较浅,根系分泌物主要富集于表层,导致土壤表层有机碳含量较高。因此,随着土壤深度的增加,有机碳含量一般会逐步降低。如三江平原湿地土壤腐殖质各组分碳含量均随土壤深度的增加而减少[23]。然而,笔者研究发现高水位区域川三蕊柳和短尖薹草群落>20~40、>40~60、>60~80 cm深度土壤有机碳和腐殖质碳含量无显著差异。沈永明等[24]的研究也发现,互花米草(Spartinaalterniflora)盐沼湿地土壤富里酸含量垂直分布差异不明显。导致不同研究之间结论存在差异的主要原因是研究区域水文条件不同[25-26]。洞庭湖为季节性淹水湿地(每年淹水时间约6个月),中下层土壤有机碳分解缓慢,这可能是不同深度土壤有机碳含量差异不显著的主要原因。
同一土壤深度下,川三蕊柳和短尖薹草2种植物群落在3种水位梯度下土壤有机碳和腐殖质碳含量均差异显著(P<0.05)。对短尖薹草群落而言,土壤有机碳含量总体上以中水位区较高。实地调查发现,中水位区短尖薹草生物量为16.14 kg·m-2,低水位和高水位区的生物量分别为12.41和6.45 kg·m-2。因此,中水位区植物生物量较高是导致短尖薹草土壤有机碳含量较高的主要原因。对川三蕊柳群落而言,植物生物量也以高水位区较低,中低水位区较高。然而,该植物群落土壤有机碳的变化规律却不同于短尖薹草,0~20 cm土壤以高水位区较高,中低水位区较低。可能原因在于湿地土壤中的碳除了来自于植物外,还有一部分来自于洪水携带的泥沙淤积。川三蕊柳冠幅宽大、枝条柔顺,能缓解水流速率,导致泥沙淤积量较高,因此高水位区川三蕊柳群落较高的泥沙淤泥量可能是表层土壤有机碳含量较高的主要原因之一。在杭州湾芦苇湿地,随着淹水深度的增加,0~20 cm土壤有机碳也逐步增加[27]。
在腐殖质各组分中,2种植物群落均以胡敏素碳含量最高,表明研究区域土壤腐殖质以胡敏素为主。在我国松嫩平原湿地区,土壤腐殖质也以胡敏素为主[10]。高湿度环境有利于土壤胡敏酸和胡敏素的形成及积累,低湿度环境有利于土壤富里酸的形成与积累[25,28]。洞庭湖属于亚热带季节性淹水湿地,调查期间地下水位为>20~60 cm,有利于土壤胡敏酸和胡敏素的积累。此外,土壤微生物种类和土壤理化性质等也会影响土壤腐殖质的形成[29]。在广西英罗港红树林湿地,淹水环境抑制了土壤腐殖化分解,腐殖酸碳含量与土壤氮磷等大量元素含量呈正相关[30-31]。张林海等[32]研究也发现,土壤胡敏素含量与土壤黏粒含量呈负相关,与土壤氮含量呈正相关关系。随着土壤含水量的提高,黏粒结合胡敏素的缩合度和芳香度增强,导致胡敏素分子结构趋于复杂,降解难度增大,从而使得土壤胡敏素含量提高[33]。
同一水位梯度下,2种植被类型之间土壤有机碳和腐殖质碳各组分差异显著。在低水位区,土壤有机碳和富里酸碳含量以川三蕊柳群落较高,而胡敏酸和胡敏素碳含量无明显差异。产生这种结果的主要原因在于低水位区泥沙淤积量较少,植物凋落物主要位于表层,因此土壤有机碳含量及组分与生物量有关。研究发现,在低水位区,川三蕊柳群落生物量高于短尖薹草,有利于土壤有机碳的积累。而低水位区土壤含水量较低,表层凋落物腐殖化过程中向富里酸转换较多,因此在土壤有机碳含量较高的短尖薹草群落土壤富里酸含量高。在中水位区,由于泥沙淤积量增加,植物凋落物会被泥沙掩埋,土壤有机碳含量的高低可能与凋落物分解过程有关。短尖薹草为草本植物,分解速率快,导致土壤有机碳、胡敏酸碳和胡敏素碳含量较高。在高水位区,土壤有机碳、胡敏酸碳和富里酸碳含量在0~20 cm深度以川三蕊柳群落较高,在>20~60 cm深度以短尖薹草群落较高,在>60~80 cm深度无明显差异。这可能是由于在高水位区,川三蕊柳群落泥沙淤积量较高,因此表层土壤有机碳、胡敏酸碳和富里酸碳含量较高。而在高水位区,表层以下土壤有机碳含量则与被泥沙掩埋的凋落物分解有关,短尖薹草较高的分解速率导致>20~60 cm土壤有机碳含量较高。可见,植物类型和水位对土壤有机碳及腐殖质碳含量具有综合影响:与短尖薹草群落相比,川三蕊柳群落能提高低水位区0~80 cm和高水位区0~20 cm土壤有机碳及腐殖质碳含量,反而降低了中水位区0~80 cm和高水位区>20~60 cm土壤有机碳及腐殖质碳含量。可见,湿地土壤碳含量受植物类型、水文特征等多种环境因子的综合影响,不能简单地以植物类型作为评估土壤碳库高低的唯一标准,而需要结合湿地水文特征,特别是水位梯度来科学评估土壤碳库。该研究主要关注土壤有机碳含量,而湿地土壤碳来源于生物固碳和泥沙沉积的外源输入碳,各来源在土壤碳库中的贡献尚不清楚,这有待结合湿地水文特征开展进一步研究。
