两个品种人参菜形态、解剖结构比较与核型分析
2019-08-29严文一谢永东仰路希王海霞贺忠群
严文一,谢永东,仰路希,陈 延,王海霞,孙 勃,贺忠群
(四川农业大学 园艺学院,四川 成都 611130)
人参菜[Talinumcrassifolium(Jacq.) Gaertn.],又名菜用土人参、绿兰菜、玉兰菜,为马齿苋科土人参属多年生热带草本植物,原产热带美洲,分布于西非、拉美、南美等热带地区[1]。在我国分布于贵州松桃、凯里、遵义、毕节、兴义、织金等地,其他地区也有少量分布[2]。人参菜具有丰富的营养价值,其可食用部位主要为嫩叶、嫩梢[3]。人参菜还具有药用价值,民间以根入药,为贵州苗族习用药材[1],有补气润肺、止咳、调经之功效,可用于治疗气虚乏力、体虚自汗、脾虚泄泻、肺痨咳血、眩晕、月经不调、乳汁稀少等。
目前,对于人参菜的研究主要集中在其栽培技术[4]、使用价值[5],对其微观结构和品种间差异比较研究基本没有。不同品种的植物在外形和显微结构上存在差异。比如空气凤梨与其他植物叶片有明显差异,其不同品种之间叶片结构也有差异[6];锦葵科花粉的形态特征在科、族、亚族和属的划分上有较大的分类学意义,从花粉形态特征角度支持将锦葵科植物划分为4个族[7];不同铁线莲属植物的花部构成上存在种间差异,柱头作为实现授粉成功的决定因素之一,与其他花部特征达成了传粉功能上的一致[8];川西高原不同山海棠的根茎解剖结构也有差异[9]。各种生物染色体的数目、结构和形态都是相对稳定的,染色体具有种的特异性,核型分析对植物种质资源的鉴定、分类、起源与进化研究具有重要意义[10]。王伟涛等[11]就不同品种梅花进行核型分析,这些指标具有一定分类学意义。本论文主要对2个人参菜品种进行形态解剖,观测2个品种的差别,以期为人参菜品种鉴别和优良品种选育提供参考依据。
1 材料与方法
1.1 材料
用于试验的2种人参菜材料由四川农业大学园艺学院设施系贺忠群教授提供。试验选取叶片、茎秆、花朵、花轴、柱头、花粉粒和蒴果作为观测材料。取材时间从人参菜开花至种子成熟。叶片为生长健壮、无病虫害植株的成熟功能叶,叶片采集方位尽量保持一致;茎秆与叶片选取相同植株;选取花瓣完全展开时候完整无损坏的柱头和花药。将所采集的样品及时带回实验室,按不同试验要求与方法进行处理。
1.2 方法
1.2.1 形态指标
花朵完全展开时,随机选取2个品种人参菜各10株,使用直尺测量植株基部到顶端的最大自然高度作为株高;从每个植株取完全展开叶片3片,使用LA-S系列植物根系分析仪扫描测定其叶片面积,winRHIZO根系扫描分析软件处理扫描结果,取其平均值作为叶片的面积;叶片厚度使用游标卡尺。选取2个品种人参菜各18株,用游标卡尺对其植株中部的茎粗进行测量;每个植株取完全展开花朵18朵,测量花的直径。待种子成熟时,随机选取2个品种人参菜各10株,每个植株取蒴果5个,称量种子千粒重(TKE)。各数据采用Microsoft Excel进行处理。
1.2.2 叶片表皮结构观测
采用表皮撕片法,对2个品种人参菜植株的叶片表皮显微结构进行观测。花朵完全展开时,取相同部位叶片中部,采用透明指甲油印迹法撕取上下表皮[12]。观测上下表皮细胞和气孔,气孔长、宽度为15个气孔平均值,气孔数量为3个视野平均值,计算气孔密度[13],数据采用Microsoft Excel进行处理。
1.2.3 花粉粒观测
在花朵完全闭合时、花朵未完全展开时、花朵完全展开时分别取样,用FAA固定液将2个品种人参菜不同时期花药样品固定,常温下,用蒸馏水清洗3次,然后将样品浸入1 mol·L-1NaOH溶液中软化8 h,用蒸馏水冲洗3次后将样品置于载玻片上,置于显微镜下观察。
1.2.4 花柱显微结构观测
在现蕾时期、初开放时期、花朵完全展开时期分别取2个品种人参菜的柱头,用FAA固定样品。常温下,用蒸馏水清洗3次后将样品浸入1 mol·L-1NaOH溶液中软化8 h左右,再用蒸馏水冲洗3次后将样品置于载玻片上,滴上数滴0.01%苯胺蓝溶液后用盖玻片盖上,约5 min后在荧光显微镜下观察花粉在花柱上的萌发部位,及花粉管生长情况[14-15],用电脑连接显微镜进行系统拍照。
1.2.5 茎横截面显微结构观测
石蜡切片法观察茎横截面[16-17]。在花朵完全展开时,将人参菜新鲜茎用FAA固定,经乙醇脱水、二甲苯透明,用石蜡包埋。切片厚度10 μm,用番红-固绿染色,用于观测2个品种人参菜茎的外形特点。
1.2.6 染色体制片与核型分析
采用根尖压片法对2种人参菜植株的染色体进行制片[18]。切取人参菜根尖约1.0 cm,放入0.002 mol·L-1的8-羟基喹啉溶液内4 ℃预处理4 h,然后在卡诺氏Ⅰ液(乙醇与冰乙酸的体积比为3∶1)、4 ℃下固定24 h,用乙醇溶液保存,再加入1 mol·L-1HCl于60 ℃水浴中解离17 min,然后采用卡宝品红染液染色制片,在显微镜下观察,找出染色体分散好的中期细胞分裂相进行拍照。
核型分析采用李懋学等[19]提出的标准,确定着丝粒位置,根据染色体的大小和形态特征对染色体核型进行配对分析。染色体的相对长度、臂比与类型按照Levan等[20]的命名法则计算,着丝粒指数和染色体相对长度系数参照李懋学等[19]的标准计算,核型不对称系数采用Arano[21]的方法计算,核型类型按照Stebbins[22]的标准划分。
2 结果与分析
2.1 形态学分析
2个品种的人参菜在外形上有较大差异。品种2具有分枝,而品种1没有;品种2株高(P<0.01)极显著大于品种1,品种2株高为83.87 cm,品种1株高为43.73 cm;品种1茎粗约为5.29 mm,极显著小于(P<0.01)品种2(8.55 mm)。与品种2相比,品种1叶片厚度大且叶片面积小,品种1叶片厚度为0.42 mm,叶面积为23.86 cm2,品种2叶片厚度为0.35 mm,叶面积为28.39 cm2。品种1叶片呈椭圆形,品种2叶片较狭长。2种人参菜花色均呈粉红色,品种2花朵极显著(P<0.01)大于品种1;2个品种的千粒重无显著差异(P>0.05)(图1,表1)。
2.2 显微结构形态
2.2.1 叶片表皮显微结构
由图2可见,2个品种人参菜上下表皮细胞均紧密排列,细胞大小形状不规则,垂周壁形状呈波浪形,但下表皮细胞较上表皮细胞更为狭长。2个品种人参菜上下表皮均有气孔,气孔由气孔、保卫细胞及副卫细胞组成。上表皮气孔比下表皮气孔大、密度小。品种1上表皮气孔明显小于品种2,但气孔密度大;品种1下表皮气孔大于品种2,但气孔密度小。2个品种上下表皮气孔的长轴方向不固定,方向随机。
2.2.2 花粉粒显微结构
2个品种人参菜的花粉粒显微结构存在较大差异。品种1的花粉粒较大,花粉粒平均直径约为42.16 μm,极显著(P<0.01)高于品种2(54.44 μm)。2个品种花粉粒均为近球形,在100倍显微镜放大观察发现,品种1人参菜花粉粒外壁无明显纹饰,而品种2花粉粒外壁纹饰可见近似正六边形紧密排列。

图1 两种人参菜形态特征Fig.1 Morphological characteristics of two Talinum crassifolium(Jacq.) Gaertn. species
表1 两种人参菜的形态特征比较
Table 1 Comparison of morphological characteristics of twoTalinumcrassifolium(Jacq.)Gaertn. species

品种Species株高Plant height/cm茎粗Stem diameter/mm叶厚度Leaf thickness/mm叶面积Leaf area/cm2花直径Flower diameter/cm千粒重1 000-seedweight/g143.73±1.81 bB5.29±0.42 bB0.42±0.03 bB23.86±2.03 bB 0.75±0.05 bB0.25±0.00 aA283.87±2.76 aA8.55±0.61 aA0.35±0.02 aA28.39±2.18 aA2.02±0.12 aA0.28±0.00 aA
同列数据后无相同大写字母表示差异极显著(P<0.01),无相同小写字母表示差异显著(P<0.05)。下同。
Data in the same column marked without the same uppercase letter indicated significant differences atP<0.01, data in the same column marked without the same lowercase letter indicated significant differences atP<0.05. The same as below.
表2 两种人参菜叶表皮气孔特征
Table 2 Stomatal characteristics of the leaves epidermis of twoTalinumcrassifolium(Jacq.)Gaertn. species

品种Species上表皮气孔Upper epidermis stomata密度Density/mm-2长度Length/μm宽度Width/μm下表皮气孔Lowr epidermis stomata密度Density/mm-2长度Length/μm宽度Width/μm138.33±2.52 aA33.18±1.25 bB16.88±1.35 bB76.19±5.50 bB26.88±2.04 aA12.99±1.02 aA233.57±4.00 bB35.66±2.51 aA19.18±1.18 aA96.91±1.15aA25.05±2.31 bA9.78±1.31 bB
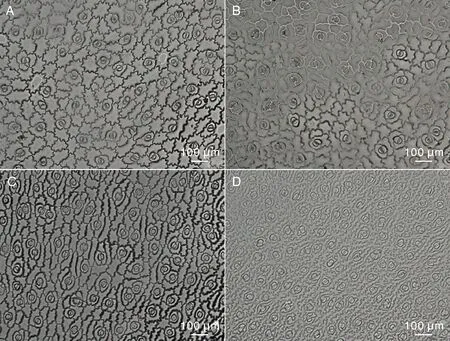
A,品种1上表皮;B,品种2上表皮;C,品种1下表皮;D,品种2下表皮。A, Upper epidermis of species 1; B, Upper epidermis of species 2; C, Lower epidermis of species 1; D, Lower epidermis of species 2.图2 两种人参菜叶表皮结构Fig.2 Leaf epidermal structure of two Talinum crassifolium(Jacq.) Gaertn. species
2.2.3 花柱的荧光显微结构
如图3所示,2种人参菜的柱头均呈三瓣羽毛状,表面有无数柱状凸起,每一瓣羽状柱头中间都有一条狭长通道。观察发现,2个品种人参菜在花瓣呈闭合状态未开放时,花柱上无花粉粒附着;花瓣完全展开时柱头上附着许多花粉粒,且花粉粒萌发出花粉管,经柱头的狭长通道到达子房。花粉管进入柱头后未见停止伸长的情况,也未见花粉粒在柱头表面萌发花粉管后没有进入柱头的情况。
2.2.4 茎的显微结构
由图4可以看出,2个品种人参菜的茎在外形上有明显区别,品种1的茎横截面近似圆形,品种2的茎横截面呈不规则形状,有4个棱角,其中3个棱角为锐角,1个棱角为钝角。2个品种人参菜茎均有表皮、皮层、维管柱,其中,皮层部分可见厚角组织和薄壁组织,品种1的茎中皮层所占整体比例比品种2小,维管柱所占比例大于品种2,且维管束排列更为紧凑;品种2的厚角组织更为明显;两者维管柱均包括维管束、髓射线和髓;维管束均呈闭合环状,与茎横截面形状贴合,且都具有韧皮部、形成层、木质部。

A,品种1花瓣完全闭合;B,品种1花瓣未全部展开;C,品种1花瓣完全展开;D,品种2花瓣完全闭合;E,品种2花瓣未全部展开;F,品种2花瓣完全展开。A, Petals of species 1were completely closed; B, Petals of species 1 were not fully unfolded; C, Petals of species 1 were fully expanded; D, Petals of species 2 were completely closed; E, Petals of species 2 were not fully unfolded; F, Petals of species 2 were fully expanded.图3 两种人参菜不同时期花柱荧光显微图像Fig.3 Fluorescence microscopic images of stigma of two Talinum crassifolium(Jacq.) Gaertn. species at different stages

A、B为品种1的茎横截面显微图像;C、D为品种2的茎横截面显微图像。1,表皮;2,薄壁组织;3,髓射线;4,韧皮部;5,形成层;6,木质部;7,髓;8,厚角组织。A and B, Stem cross section of species 1; C and D, Stem cross section of species 2. 1, Epidermis; 2, Parenchyma; 3, Pith ray; 4, Phloem; 5, Cambium; 6, Xylem; 7, Pith; 8, Collenchyma.图4 两种人参菜茎的显微结构图像Fig.4 Microscopic structure images of stem of two Talinum crassifolium(Jacq.) Gaertn. species
2.3 核型分析
2.3.1 两种人参菜染色体数目形态比较
2种人参菜的染色体如图5所示:品种1的染色体数目是40条,可配对成20对同源染色体,为二倍体,染色体基数x=20;品种2的染色体数目是24条,可以配对成12对同源染色体,为二倍体,染色体基数x=12。
2.3.2 两种人参菜的核型分析
核型分析结果(表3)显示:品种1核型公式为2n=2x=38m+2M(n表示物种配子体的染色体个数;x表示该1个染色体组中染色体数,即染色体基数),38条为中部着丝粒染色体(m),2条为正中部着丝粒染色体(M);品种2的核型公式为2n=2x=18m+6sm,其中18条为中部着丝粒染色体(m),6条为近中部着丝粒染色体(sm)。品种1的染色体相对长度组成为8L+14M2+12M1+6S,包含4对长染色体(L),7对中长染色体(M2),6对中短染色体(M1)和3对短染色体(S);品种2的染色体相对长度组成为6L+4M2+8M1+6S,包含3对长染色体(L),2对中长染色体(M2),4对中短染色体(M1)和3对短染色体(S)。由表3和图6可知,2个人参菜的相对长度和臂比都存在明显差异,品种1的染色体相对长度为2.52%~8.46%,染色体相对长度最长与最短的比值为3.36,且没有臂比值大于2∶1的染色体;品种2的染色体相对长度为2.24%~6.56%,染色体相对长度最长与最短的比值为2.93,且臂比值大于2∶1的染色体百分比为16.67%。
3 结论与讨论
2个品种人参菜在外形上差异大,叶片、花粉粒和茎显微结构差异较大,柱头和花粉管萌发伸长差异不大,这是因为2个品种人参菜同属不同种,且生长环境也不同造成的。品种2更粗壮,叶片厚度、叶片面积、茎粗、花直径均极显著大于品种1。高大的品种2相对于矮小的品种1,周围环境空气流动大,光照更足,更易进行蒸腾作用,因此品种2叶片增厚可以减小水分蒸腾[23]。
2个品种叶片表皮细胞排列紧密,形状大小不规则,垂周壁形状均呈波浪状,下表皮细胞比上表皮细胞更为狭长;上下表皮气孔长轴方向随机,与膝柄木一样[24],气孔具有1个细胞的间距,这种单细胞间距与Bergmann等[25]的结论一致。姚兆华等[26]提出气孔的大小与密度具有分类学意义,尤其是气孔大小种间差异明显。2个品种人参菜上表皮气孔密度小于下表皮,品种1叶片上表皮气孔小于品种2,下表皮气孔大于品种2。可能是由于品种1植株矮小,被其他高大植物遮挡,气孔密度大可以增强叶片空气交换能力。
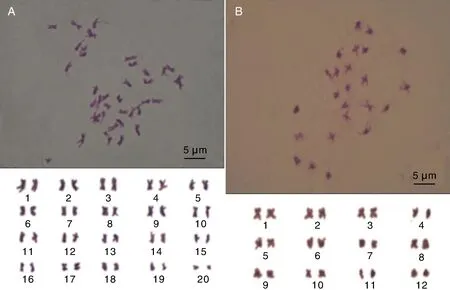
A,品种1;B,品种2。A, Species 1; B, Species 2.图5 两种人参菜染色体数目形态Fig.5 Chromosome number and morphology of two Talinum crassifolium (Jacq.) Gaertn. species
表3 两种人参菜染色体核型参数
Table 3 Karyotype parameters of twoTalinumcrassifolium(Jacq.) Gaertn. species

品种Species染色体编号Chromosomenumber相对长度(S+L=T)Relative length/%相对长度系数Relative lengthcoefficient着丝粒指数Centromereindex臂比Armratio/%类型Type114.03+4.43=8.461.410.481.10m23.83+4.03=7.861.310.491.05m33.09+4.76=7.851.310.391.54m43.51+4.28=7.791.300.451.22m53.29+3.58=6.871.140.471.09m63.26+3.34=6.591.100.491.02m73.26+3.26=6.521.090.501.00M83.01+3.51=6.521.090.461.17m92.86+3.58=6.441.070.441.25m103.01+3.41=6.421.070.471.13m112.76+3.29=6.051.010.461.19m122.57+3.29=5.850.970.441.28m132.76+3.04=5.800.970.481.10m142.51+3.26=5.760.960.431.30m152.51+2.66=5.170.860.491.06m162.14+2.77=4.900.820.441.30m172.11+2.80=4.900.820.431.33m181.77+2.76=4.530.750.391.56m191.26+2.06=3.320.550.381.64m201.24+1.28=2.520.420.491.03m212.52+4.04=6.561.510.381.60m22.51+3.53=6.041.370.421.41m32.76+3.27=6.031.370.461.18m42.24+2.29=4.531.030.501.02m51.27+3.26=4.521.030.282.57sm61.58+2.76=4.340.990.361.75sm71.58+2.63=4.210.960.381.66m81.51+2.51=4.020.920.381.66m91.11+2.76=3.870.880.292.50sm101.52+1.77=3.290.750.461.16m111.51+1.52=3.030.690.501.01m121.07+1.18=2.240.510.481.10m
S,短臂;L,长臂;T,全长;m,中部着丝粒染色体;M,正中部着丝粒染色体;sm,近中部着丝粒染色体。
S, Short arm; L, Long arm; T, Total length; m, Metacentric chromosome; M, Choromosome structure of median poin; sm, Submetacentric chromosome.
花粉形态的演化主要表现在外壁纹饰和萌发孔上[27],花粉形态特征受基因型控制而不受外界条件影响,是探讨植物起源、演化、亲缘关系的重要特征之一,可根据花粉形态进行植物品种间的鉴定、分类。2个品种的花粉粒形状无较大差异,都是近球形。但在大小和外壁纹饰上有极大差异,品种2花粉粒比品种1大,并且其外壁有明显可分辨的近正六边形紧密排列纹饰,品种1表面无明显纹饰。2个品种的花柱形态相似,均呈三瓣羽毛状,表面有无数柱状凸起,每一瓣羽状柱头中间都有一条狭长通道。2个品种人参菜虽然是雌雄同花,但由于花瓣完全闭合时未见花粉粒附着柱头,在花瓣完全展开时才有花粉粒附着柱头,因此2个品种人参菜可能不仅进行自花授粉,还通过风、昆虫等媒介进行异花授粉。

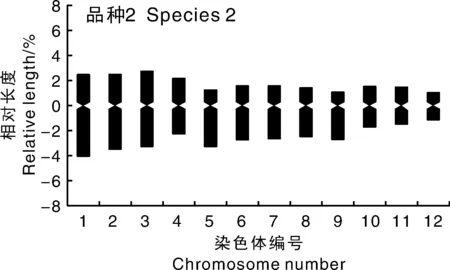
图6 两种人参菜染色体核型模式图Fig.6 Karyotype pattern of chromosomes of two Talinum crassifolium (Jacq.) Gaertn. species
2个品种人参菜茎的组成部分无较大差别。品种1的茎中皮层所占比例比品种2小,维管柱所占比例大于品种2,排列更为紧凑。品种2的厚角组织更为明显,使得品种2的茎秆更为坚挺,起到很好的机械支撑作用。
品种2染色体数目与陈瑞阳等[28]的测定结果一致,但是染色体的具体核型参数有差异,这说明品种2的染色体的数目比较稳定,但染色体大小可能受环境等因素影响。品种1的染色体数目与陈瑞阳等[28]的报道不一致;美国得克萨斯州的一个马齿苋科物种(Claytoniavirginica)居群中就发现染色体数目可从2n=12变化到2n=42[29],品种2和品种1的核型公式与相对对长度系数组成差别都比较明显,在其他种的染色体研究中也有不同品种染色核型差异的报道[30-32]。根据Stebbins的不对称核型分析的分类标准,品种1的核型属于1B型,核型不对称系数为54.50%;品种2的核型属于2B型,核型不对称系数为59.37%。核型不对称系数越接近50 %,核型的对称程度越高,进化程度越原始[22],品种2和品种1的核型不对称系数都比较低,在系统演化中具有较强的原始性。综上所述,可能染色体核型不同使2个人参菜植株性状表现出差异,品种1和品种2可能是同种不同属的2种处于半野生状态的植株。
