Prolonged high-fat-diet feeding promotes non-alcoholic fatty liver disease and alters gut microbiota in mice
2019-08-29KandyVelzquezReillyEnosJackieBaderAlexanderSougiannisMeredithCarsonIouliaChatzistamouJamesCarsonPrakashNagarkattiMitziNagarkattiAngelaMurphy
Kandy T Velázquez,Reilly T Enos,Jackie E Bader,Alexander T Sougiannis,Meredith S Carson,Ioulia Chatzistamou,James A Carson,Prakash Nagarkatti,Mitzi Nagarkatti,E Angela Murphy
Kandy T Velázquez,Reilly T Enos,Jackie E Bader,Alexander T Sougiannis,Meredith S Carson,loulia Chatzistamou,James A Carson,Prakash Nagarkatti,Mitzi Nagarkatti,E Angela Murphy,Department of Pathology,Microbiology and Immunology,School of Medicine,University of South Carolina,Columbia,SC 29209,United States
James A Carson,College of Health Professions,University of Tennessee Health Sciences Center,Memphis,TN 38163,United States
Abstract
Key words: High-fat diet;Obesity;Non-alcoholic fatty liver disease;Gut microbiome;Endoplasmic reticulum stress;Inflammation;Fibrosis
INTRODUCTION
Advanced stages of non-alcoholic fatty liver disease (NAFLD) can be classified as a form of chronic hepatitis.An estimated 25% of individuals are affected globally[1].In the United States alone,approximately 95 million adults have NAFLD and the prevalence has continued to rise[2].In fact,it is now considered the most common cause of liver disorder in the United States and other Western industrialized countries.NAFLD exists as a spectrum and is best categorized histologically.Characteristic features include steatosis,inflammation,hepatocellular ballooning,and fibrosis[3].NAFLD can be further classified as non-alcoholic fatty liver (NAFL) and non-alcoholic steatohepatitis (NASH)[3].NAFL is a less severe form and is defined as the presence of steatosis with no evidence of hepatocellular injury in the form of ballooning of the hepatocytes[3].NASH,on the other hand,is characterized by the presence of hepatic steatosis and inflammation with hepatocyte injury (ballooning)with or without fibrosis[3].These characteristics of NAFLD can facilitate the risk of further disease progression[1].For example,NAFLD has been linked to the progression and development of hepatocellular carcinoma (HCC),the major histological subtype of liver cancer[1].Similarly,it has been associated with metabolic disorders as up to 75% of individuals with NAFLD have been reported to have type II diabetes[1].Thus,understanding the risk factors for NAFLD is of critical public health importance.
Obesity is a well-characterized risk factor for the development of NAFLD.Although it is well known that obesity can be prevented through healthy dietary habits and physical activity[4,5],interventions in a clinical setting have largely been unsuccessful,especially in the long-term[6,7].Thus,recent research has focused on understanding the pathways driving the pathologic processes associated with obesityinduced NAFLD so that therapeutic targets can be identified.Animal models are critical to this mission and have greatly enhanced our understanding of NAFLD development.Although multiple animal models of NAFLD exist,high-fat-diet (HFD)administration has been a widely used model[8].However,a limitation of this approach is that HFD administration to mice does not appear to induce significant NAFLD progression (i.e.,liver cell death,inflammation,or fibrosis) despite reproducibly inciting obesity,the metabolic syndrome,and hepatic steatosis[8].A potential explanation for this phenomenon is that the duration of HFD feedings is not long enough to produce significant NASH even when diet treatment is administered for six months.Therefore,a longer duration of HFD administration may be necessary to recapitulate the pathology seen in the human condition.
The gut microbiome has recently emerged as a culprit in the development of chronic diseases,such as,obesity[9,10],diabetes[11,12],liver disease[13,14],and cancer[15].In the case of NAFLD,the location of the portal vein allows easy access for bacteria and microbial-derived products to translocate from the gut to the liver[16].In support of this hypothesis,studies conducted in obese humans with NAFLD revealed gut dysbiosis when compared to healthy humans[17,18].Ramanet al[17]reported an enrichment ofLactobacillusspecies and various microbes from theFirmicutesphylum in obese humans with NAFLD.Meanwhile,Wonget al[18],observed that NASH patients had lower fecal abundance ofFirmicutes.Although studies examining the link between human gut microbiota and liver diseases have advanced our understanding of this relationship,preclinical models mimicking gut dysbiosis in NAFLD are still lacking[19].
We sought to examine the effects of chronic HFD feeding on NAFLD in mice.C57BL/6mice were used given their susceptibility to HFD-induced obesity.We utilized a diet consisting of 60% fat,20% protein,and 20% carbohydrate,which was fed to the mice for a period of 80 wk-a protocol designed to mimic lifetime consumption of a diet high in fat.Our analysis focused largely on liver pathology,fibrosis,inflammation,and endoplasmic reticulum (ER) stress.We also measured metabolic outcomes and characterized fecal microbiota using 16S rRNA sequencing.Our data indicate that chronic HFD consumption does result in significant NAFLD and gut-bacterial dysbiosis.Specifically,we report a significant increase in steatosis,inflammation,cell injury,fibrosis,and ER stress,which was associated with increases in theActinobacteriaandFirmicutesphylum and decreases in theBacteriodetesandTenericutesphylum.
MATERIAL AND METHODS
Animals and diet
All procedures involving animals were reviewed and approved by the Institutional Animal Care and Use Committee at the University of South Carolina (animal protocol number,2044-100482-093011).A total of 53C57BL/6male mice were obtained from Jackson Laboratories.Mice (3-5 per cage,wood bedding with nesting material) were kept in 12:12 h dark/light cycle,in a humidity and temperature control room,andad libitumaccess to food and water.Animal handling and experiments were performed to minimize pain and discomfort.At 10 wk of age,male mice were randomly assigned to one of two diets for the duration of 80 wk:Low-fat diet (old-LFD,n= 15) from Harlan Teklad Rodent Diet,no 8604 (14% Fat,54% carbohydrate,32% protein) or High-Fat Diet (Old-HFD,n= 18) from Research Diets,D12492 (60% Fat,20%carbohydrate,20% protein).An additional group of male mice (10 wk of age) was maintained on the LFD (Young-LFD,n= 20) but for a shorter duration (6 wk) to distinguish between age-dependent and age-independent effects of obesity on metabolic,molecular,and histological measures.All three groups of mice were euthanized (overdose of isoflurane) the same day,fat depots (epididymal,retroperitoneal,and mesentery),liver,and spleen were removed and weighed.The livers were collected and stored at -80 °C or fixed in 4% paraformaldehyde for further analysis.
Metabolic measurements and assays
A day prior to euthanasia,ten mice per each group were fasted for five hours (light cycle).Blood collection was carried out in conscious animals,the tip of the tail was cut with scissors and heparinized capillary tubes (0.12 cm diameter,7.5 cm length) were used to collect 70 μL of blood from the tail vein for the measurement of glucose and insulin.Blood glucose was assessed using a glucometer (Bayer Counter,New Jersey,United States) and plasma insulin was determined using a mouse ELISA assay from Mercodia (Uppsala,Sweden).Insulin resistance was calculated by HOMA index using the following equation IR = (insulin uU/mL) (glucose mmol/L) /22.5.
Staining
Liver,colon,and adipose tissue were fixed in 4% paraformaldehyde,paraffinembedded,sectioned,and then stained with hematoxylin eosin (HE).Picro-sirius red stain kit (Cat ab150681,abcam,Cambridge,MA,United States) was used according to the manufacturer’s instructions to stain the liver for histological evaluation of fibrosis.For Oil red O staining,frozen liver tissues were cut (10 μm) using a cryostat (Leica Biosystems,Nussloch,Germany) and staining was performed as previously described[20].
Histopathology
Histological scoring system for NAFLD was achieved based on HE and picro-sirius red staining in the liver of Young-LFD (n= 6),Old-LFD (n= 10),and Old-HFD (n=12) mice as previously described[21].A certified pathologist (I.C.) blindly evaluated the histological findings of steatosis (0-3),portal and lobular inflammation (0-3),cell injury (0-2),and fibrosis (0-4) in the liver sections.NAFLD Activity Score (NAS) was calculated by adding the unweighted scores for steatosis,lobular inflammation,cell injury (0-8).
Western blotting
Briefly,liver was homogenized in Mueller Buffer containing a protease inhibitor cocktail (Sigma Aldrich,St.Louis,MO,United States).Total protein concentrations were determined by the Bradford method.Equal amounts of crude protein homogenates (20 μg) were fractioned on hand-casted SDS-polyacrylamide gels and electrophoretically transferred to a PVDF membrane using a Royal Genie Blotter(IDEA Scientific,Minneapolis,MN,United States).Membranes were stained with a Ponceau S solution in order to verify equal protein loading and transfer efficiency.Western blots were performed using primary antibodies from Cell Signaling(Danvers,MA,United States) (phosphorylated-IRE1α,IRE1α,XBP1,phosphorylated-EIF2α,EIF2α,phosphorylated-Jnk,Jnk,CHOP,β-actin,phosphorylated-NFκB,NFκB),AbD Serotec Raleigh,NC (F4/80),and Novus Biologicals Littleton,CO(phosphorylated-IRE1α).
Quantitative real-time PCR
Gene expression in liver and colon tissue was performed in duplicate after RNA isolation with trizol reagent.The following Taqman gene expression assays from Applied Biosystems were used:Monocyte chemoattractant protein-1 (MCP-1),interleukin 10 (IL-10),interleukin 17-alpha (IL17α),interleukin 6 (IL-6),forkhead box P3 (Foxp3),or tumor necrosis factor alpha (TNFα).All primers were normalized to 18s rRNA.
Microbiome analysis
Before sacrifice,mice were individually placed in autoclaved cages with no bedding for the collection of three or more fecal pellets per mouse.Fecal pellets were frozen immediately after collection and stored at -80 °C.Isolation and concentration of microbial DNA was achieved using the QIAamp Fast DNA Stool Mini Kit (cat number:51604,QIAGEN) and a nanophotometer Pearl,respectively.Characterization of the fecal microbiota via 16S rRNA sequencing was performed via the amplification of the 16S rRNA V3 and V4 hypervariable regions.We used 16S V3 314F forward and V4 805R reverse primers,respectively (5′TCGTCGGCAGCGTCAGATGTGTATA AGAGACAGCCTACGGGNGGCWGCAG3′&5′GTCTCGTGGGCTCGGAG ATGTGTATAAGAGACAG GACTACHVGG GTATCTAATCC3′) with added Ilumina adapter overhang nucleotide sequences[22].The PCR conditions used were 3 min at 95°C followed by 25 cycles of 30 s at 95 °C,30 sat 55 °C,30 s at 72 °C,and a final extension at 72°C for 5 min.Each reaction mixture (25 μL) contained 50 ng of genomic DNA,0.5 μL of amplicon PCR forward primer (0.2 μmol/L),0.5 μl of amplicon PCR reverse primer (0.2 μmol/L) and 12.5 μl of 2× KAPA Hifi Hot Start Ready Mix.Each reaction was cleaned up using Agencourt AMPure XP beads.Attachment of dual indices and Illumina sequencing adapters was performed using 5 μL of amplicon PCR product DNA,5 μL of Illumina Nextera XT Index Primer 1 (N7xx),5 μL of Nextera XT Index Primer 2 (S5xx),25 μL of 2× KAPA HiFi Hot Start Ready Mix,and 10 μLμL of PCR-grade water.Amplification was carried out under the following conditions:3 min at 95 °C,followed by 8 cycles of 30 s at 95 °C,30 s at 55 °C,and 30 s at 72 °C,and a final extension at 72 °C for 5 min.Constructed 16S metagenomic libraries were purified with Agencourt AM Pure XP beads and quantified with Quant-iTPicoGreen.Library quality control and average size distribution were determined using an Agilent Technologies 2100 Bioanalyzer.Libraries were normalized and pooled to 40 nmol/L based on quantified values.Pooled samples were denatured and diluted to a final concentration of 6 pmol/L with a 30% PhiX (Illumina) control.Amplicons were subjected to pyrosequencing using the MiSeq Reagent Kit V3 in the Illumina MiSeq System.The online 16S analysis software from NIH was used to analyze sequencing data collected on the Illumina Miseq.FASTQ sequences were uploaded to Nephele and the 16S metagenomics application was executed.The groups of related DNA sequences were assigned to operational taxonomic units (OTUs),and output files were analyzed to determine gut microbial composition.
Statistical analysis
Data from this experiment were analyzed using commercially available statistical software,Prism 5 (GraphPad Software).The statistician,Dr.Bo Cai,from the Department Epidemiology and Biostatistics at the University of South Carolina reviewed the statistical methods of this study.Morphometric measurements,metabolic assays,western blot analysis,and gene expression results were analyzed using a one-way ANOVA followed by Newman-Keuls post hoc test.Kruskal-Wallis test followed by a Dunn’s Post-Hoc was used to assess differences in histopathology.A two-tailed Student’s t-test and a Mann-WhitneyUtest were used to determine differences in microbiota phylum,family,and genus between Old-LFD and Old-HFD mice.Data that did not pass Barlett’s test for equal variances was log-transformed and then re-analyzed.Data are presented as the mean ± SE or median with interquartile range.The level of significance was set atP< 0.05.
RESULTS
Morphometric and metabolic analysis of diet-induced NAFLD
To evaluate whether prolonged HFD feeding leads to morphometric and metabolic changes,we determined the mass of three visceral fat depots (epididymal,kidney,and mesentery) and evaluated basal glucose metabolism.As expected,20 mo of HFD feedings augmented body weight (Figure 1A) in Old-HFD mice when compared to Young-LFD and Old-LFD mice (P< 0.05).This increase in body weight was in part due to the significant expansion of epididymal (Figure 1B),kidney (Figure 1C),and mesentery (Figure 1D) fat pads (P< 0.05) as well as liver (Figure 1E) and spleen(Figure 1F) tissues.In addition,Old-LFD mice showed a higher body weight and epididymal fat accumulation than Young-LFD mice.Figure 1G,shows a representative histological image of epididymal fat tissue stained with HE;we can observe larger adipocytes surrounded by infiltrated immune cell in Old-HFD mice compared to Old-LFD and Young-LFD mice.
We next addressed whether Old-HFD mice were insulin resistant.As anticipated,Old-HFD mice showed significantly elevated fasting blood glucose (Figure 2A) and insulin (Figure 2B) when compared to Young-LFD and Old-LFD mice (P< 0.05).These results were consistent with the HOMA index in which Old-HFD mice exhibited insulin resistance relative to Young-LFD and Old-LFD mice (P< 0.05)(Figure 2C).No differences in glucose,insulin,and HOMA index were observed between Old-LFD and Young-LFD mice even though Old-LFD mice displayed an increase in body weight and epididymal fat mass.
Histopathological assessment of diet-induced NAFLD
In order to evaluate whether prolonged HFD promotes NAFLD,we performed HE,picro-sirius red,and Oil Red O stains in the liver tissue of Young-LFD,Old-LFD,and Old-HFD mice (Figure 3A-C).The analysis of the specimens showed that livers of the Young-LFD mice had minimal focal inflammation,minimal perisinusoidal fibrosis,and no signs of steatosis (NAS score 0-1) (Figure 3D-J).Old-LFD mice displayed mild focal inflammation with focal steatosis,and perisinusoidal fibrosis (NAS score 1-2).In the case of Old-HFD mice,we observed extensive steatosis,portal and lobular inflammation with cell injury (ballooning degeneration of the hepatocytes),and evident fibrosis (NAS score 5-6).We next confirmed fat accumulation in the liver with Oil Red O staining (Figure 3C).Overall,Oil Red O staining provided evidence of macrovesicular accumulation of triglycerides in the hepatocytes of Old-HFD mice when compared to Young-LFD and Old-LFD groups.
Examination of inflammatory markers in the liver tissue of mice following prolonged HFD feedings
Since inflammation is one of the hallmarks of obesity and NAFLD,we next addressed the question of whether prolonged HFD consumption increases liver inflammation.To assess this,we examined gene expression of MCP-1,IL-6,and TNF-α and the protein concentration of F4/80 and p-NFκB (Figure 4A-E).MCP-1 was significantly elevated in Old-HFD and Old-LFD mice when compared to Young-LFD mice (P<0 .05).Interestingly,F4/80 was significantly decreased with age with a further decrease observed with HFD feeding (P< 0.05).No significant changes were observed in the gene expression of IL-6 and TNFα or the protein expression of p-NFκB.

Figure1 Morphometric characterization of mice subjected to prolonged high-fat diet-feeding.
Effects of prolonged HFD feeding on ER stress in the liver
Previous studies have reported that ER stress is involved in the development and progression of steatosis[23-25].Therefore,we next investigated the signaling pathways activated by ER stress,such as,binding immunoglobulin (Bip),inositol-requiring enzyme-1 (IRE1α),X-box-binding protein-1 (XBP1s),eukaryotic translation initiation factor 2α (EIF2α),c-Jun-N-terminal kinase (Jnk),and C/EBP-homologous protein(CHOP).There was no change in Bip across the groups (Figure 5A).However,both of the aged groups showed a significant reduction in phosphorylated IRE1α (Figure 5B)and XBP1s (Figure 5C) when compared to the Young-LFD mice (P< 0.05) and there was a further reduction in pIRE1α with HFD feeding (Old-LFD versus Old-HFD) (P<0.05).For phosphorylated EIF2α protein expression,there was an elevation in the Old-LFD group compared to Young-LFD and Old-HFD (Figure 5D) (P< 0.05).Phosphorylated Jnk was increased in both of the aged groups and there was a further increase with HFD feeding (Figure 5E) (P< 0.05).Only the Old-HFD group exhibited an increase in CHOP when compared to Old-LFD and Young-LFD (Figure 5F) (P<0.05).
Effects of prolonged HFD feeding on colon inflammation and the gut microbiome profile
To examine whether prolonged HFD feeding produced an inflammatory environment in the colon and/or disrupted the gut microbiome,we analyzed gene expression of inflammatory markers in the colonic tissue and fecal microbial DNA using pyrosequencing.In this study,we did not assess the bacterial composition of the Young-LFD mice,because short-term manipulation of diet is transient[26].A significant increase in IL-6 was observed with age and there was a further increase with HFD feedings (Figure 6C) (P< 0.05).Foxp3 was increased in Old-HFD mice when compared to Young-LFD and Old-LFD mice (P< 0.05) (Figure 6C and E).However,we found no changes in MCP-1,IL-10,or IL-17α (Figure 6AB,and D) between any of the groups.Representative histological images of the colon tissue in Figure 6F shows similar colonic morphology between groups.
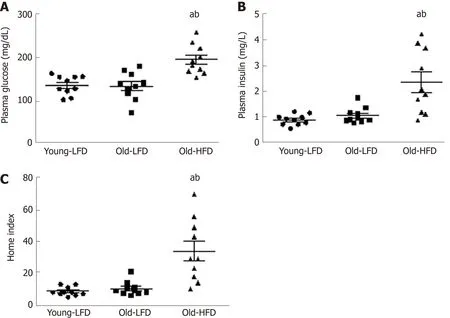
Figure2 Metabolic characterization of mice subjected to prolonged high-fat diet-feeding.
In Figure 7,we observed similar species richness at the taxonomic level (Figure 7A).However,principal coordinate (PC) analysis based on OTU showed distinct gut microbiota composition between Old-LFD and Old-HFD mice (Figure 7B).FirmicutesandBacteriodeteswere the most predominant phyla in both groups,comprising of 61%and 32% of gut microbiota in Old-LFD mice and 73% and 21% in Old-HFD mice,respectively (Figure 8A and B).Consistently,the ratio ofFirmicutestoBacteriodetesin Old-HFD mice was altered to favorFirmicuteswhen compared with Old-LFD (Figure 8C,P< 0.05).The next two prevalent phylum includedTenericutesandVerrucumicrobiawith 5% and 0.2% of microbiomes in Old-LFD mice and 0.8% and 3.4% in Old-HFD mice,respectively.Prolonged HFD feeding significantly increased the abundance ofActinobacteriaandFirmicutesand decreased the abundance ofBacteriodetesandTenericuteswhen compared to Old-LFD mice (P< 0.05).In figure 8D,a positive correlation was observed between body weight andFirmicutesabundance(P< 0.05),while a negative correlation was observed between body weight andBacteriodetesabundance (P< 0.05).At the genus level (Figure 9),we observed a significant increase in the abundance ofAdercreutzia (Phylum-Actinobacteria),Coprococcus (Phylum-Firmicutes),Dorea (Phylum-Firmicutes),andRuminococcus(Phylum-Firmicutes)in Old-HFD mice when compared to Old-LFD mice (P< 0.05).Further,Old-HFD mice showed a decrease in the abundance ofTuricibacter (Phylum-Firmicutes)andAnaeroplasma (Phylum-Tenericutes)(P< 0.05).
DISCUSSION
In humans,chronic HFD consumption is associated with systemic inflammation,obesity,metabolic dysfunction,NAFLD,and an altered gut microbial profile[27-29].However,animal studies using standard HFD paradigms (16 wk feeding) which mimic obesity,metabolic disorders,and gut microbiota disruption have failed to produce consistent NAFLD pathology[8,30]limiting our advancement of mechanistic insights and the development of therapeutic strategies.Therefore,the present study utilized a prolonged HFD feeding (80 wk) to examine NAFLD evolvement.Given that the progression of steatosis to NASH has been linked with changes in inflammation,ER stress[23],and fecal bacterial composition[17],we sought to examine these outcomes in our pre-clinical diet-induced obesity model.In general,we found that prolonged HFD feeding promotes obesity,insulin resistance,ER stress,alterations in gut bacterial composition,and NAFLD in male C57BL/6 mice.

Figure4 lnflammatory signaling in the liver tissue of mice subjected to prolonged high-fat diet-feeding.
In order to determine if lifetime consumption of a HFD can reliably reproduce NAFLD,we fed mice for a period of 80 wk with a 60% (total kcals) fat diet.NAFLD with 14% kcal from fat and an equal amount of choline as the HFD was used to compare disease outcomes independent of choline deficiency.We controlled for choline given that choline deficient diets have been widely used to induce NAFLD/NASH in rodents[31,32],despite the fact that these diets do not show physiological characteristics involved in the progression of NAFLD/NASH[33].Consistent with our previous studies[21,34-36],we report that HFD consumption increased morphometric parameters (body weight,liver mass,fat pads,and spleen mass) and fasting blood glucose and insulin.Histopathological analysis of the liver indicated that prolonged HFD feeding produced extensive steatosis with portal and lobular inflammation and parenchymal fibrosis in all mice in the Old-HFD group(NAS ≥ 5).Similar to our findings,others have reported metabolic disturbances and NAFLD pathology using an extended period of HFD feedings[37,38].For example,VanSaunet al[37]reported pericellular fibrosis,advanced stage of fibrosis,and NASH after 36,56,and 80 wk of HFD,respectively.In addition,they observed non-invasive dysplastic tumors in seven of eight mice fed HFD for 36-80 wk.Interestingly,we also observed non-invasive focal masses in the liver of Old-HFD mice.Young-LFD and Old-LFD mice had an average NAS score of 1,which is not considered diagnostic of steatohepatitis.

Figure5 Endoplasmic reticulum stress signaling in the liver tissue of mice subjected to prolonged high-fat diet-feeding.

Figure6 lnflammatory signaling in the colon tissue of mice subjected to prolonged high-fat diet-feeding.
Other HFD feeding regimes have been used to mimic the NAFLD condition.However,variability in the degree of NAFLD pathology has been a limitation of these models.For example,the use of the Amylin liver NASH model (AMLN) diet,which is based on a high content of fructose,cholesterol,and trans-fat (partially hydrogenated vegetable oil),has been shown to induce NAFLD stage heterogeneity in mice when the duration of feeding is between 12-30 wk[39-41].However,one study reported that only twenty percent of mice develop liver fibrosis when fed an AMLN diet for a duration of 12 wk[40].Another study reported that key hallmarks of NASH(macrovesicular steatosis and high levels of aspartate aminotransferase) are detected as early as 26 wk after AMLN diet consumption in Lepob/oband C57BL6 mice[41].Nonetheless,not all C57BL/6 mice developed NASH when mice were maintained on an AMLN diet for 30 wk[39].Thus,prolonging the duration of HFD consumption,as in the current study,may be enough to promote a more homogeneous pathology of NAFLD.

Figure7 Gut microbial composition of mice subjected to prolonged high-fat diet-feeding.
The role of pro-inflammatory cytokines in obesity and NAFLD remains unclear.Human studies investigating the action of plasma TNF-α has shown positive or no correlation with insulin resistance in NAFLD and obese patients[42,43].In the case of IL-6,a more clear association has been stipulated between elevated plasma IL-6,insulin resistance,and NAFLD progression[44,45].However,there is no clinical evidence that blocking IL-6 may serve as a treatment for NAFLD and/or metabolic diseases.Additionally,elevated levels of the chemokine MCP-1 also has been observed in NASH and NAFLD patients,which has been correlated with liver fat content[46-48].Our current findings show no differences in the gene expression of TNFα and IL-6 in mouse liver with NASH pathology.These data coincide with no changes in protein expression of NFκB,a transcription factor involved in inflammatory signaling pathways.Contrary to our hypothesis,a reduction in F4/80 protein expression,a pan macrophage and Kupffer cells marker,was observed in aged mice.This decrease in F4/80 may be a result of aging,as macrophage immune function declines in senescence[49,50].Old-HFD mice showed an even more significant drop in F4/80 than Old-LFD mice,which may be an indication of a depressed immune system related to NAFLD.Interestingly,the gene expression of MCP-1,a major recruiter of macrophages was increased in the liver of both Old-LFD and Old-HFD mice.Given that there was no difference between Old-HFD and Old-LFD groups we believe that this effect is due to aging and not obesity nor NAFLD and likely a compensatory response to the decrease in F4/80 seen in aged mice.It is important to mention that even though we did not find significant changes in the gene expression of TNFα and IL-6,we did observe significant lobular inflammation in the liver of Old-HFD mice,histologically.
ER stress is associated with obesity,insulin resistance,and NAFLD[23-25,51].It is believed that ER stress can lead to hepatic steatosis by altering lipid metabolism[52].The ER stress response is regulated by three transmembrane proteins:ATF6,IRE1α,and PKR-like ER kinase (PERK),the molecular chaperone,BiP,and their downstream signaling cascades,including EIF2α (PERK pathway) and XBP1 activation (IRE1α pathway),which attempt to bring homeostasis back to the cell under stressful conditions.However,if this is not achieved,chronic ER stress ensues eventually leading to cell death via Jnk and CHOP signaling[53]and downregulation of the IRE1α-XBP1 axis[54].A rapid response to acute ER stress is a significant increase in the production of molecular chaperones,such as BiP[54].We found no differences in the protein expression across groups with respect to BiP;however,we did find a significant decrease in p-IRE1α and XBP1s in both of the Old groups suggesting that the livers of these mice were experiencing a certain degree of chronic ER stress.Furthermore,the fact that the Old-HFD mice displayed increases in both p-JNK and CHOP protein expression compared to all other groups provides evidence that the chronic consumption of the HFD led to a more advanced stage of chronic ER stress than the Old LFD-treated mice.This is corroborated by the significant increase in p-EIF2α displayed by the Old-LFD mice and not the Old-HFD mice,as activation of EIF2α,which inhibits protein translation,typically occurs during the earlier stages of chronic ER stress[55].In fact,Choiet al[56]demonstrated that inhibition of EIF2α phosphorylation exacerbates macro-vesicular steatosis,leukocyte infiltration,and fibrosis in mice.Thus,the increased EIF2α phosphorylation displayed by the Old-LFD mice may be a protective mechanism against the more advance stages of NAFLD as displayed by the Old-HFD mice.

Figure8 Relative abundance of gut microbial at the phylum level of mice subjected to prolonged high-fat diet-feeding.
We also sought to investigate colonic inflammatory cytokines involved in the pathology of colitis due to the co-existence of NAFLD with inflammatory bowel disease (IBD)[57-61].We observed an increase in IL-6 and FoxP3 in the colon tissue of Old-HFD mice,but no changes were observed in MCP-1,IL-10,and IL-17α between groups.Our findings of an increase in IL-6 is consistent with Jianget al[62],that reported an increase in IL-6 mRNA expression in the intestinal mucosa of NAFLD patients when compared to healthy subjects.Since systemic inflammation including IL-6 and TNFα has been involved in both NAFLD and IBS in humans[57-61],we cannot discard that systemic inflammation and/or gut permeability may play an important role in the development of these disorders.However,we did not measure systemic cytokines nor intestinal permeability in our study.Thus,whether prolonged HFD consumption for 80 wk,as in the current study,exhibits these characteristics remains to be determined.The increase in FoxP3 expression is likely a consequence of the chronic low-grade inflammation in the colon and may explain,in part,the absence of a difference in colon histology.

Figure9 Relative abundance of gut microbial at the genus level of mice subjected to prolonged high-fat diet-feeding.
Studies have reported that the taxonomic composition of gut microbiota in obese individuals indicates a higher proportion ofFirmicutesand a lower amount ofBacteroidetes[14,19].Interestingly,these differences in the ratio ofFirmicutesandBacteroidetesat the phylum level has been not observed in obese NAFLD patients when compared with age match healthy control subjects[17,18,62-68].In fact,most of the gut microbiota associated changes in NAFLD and NASH occur at the family and the genus level (Table 1).In general,there is not consistent evidence of a specific family or genus involved in NAFLD/NASH.This could be due,in part,to discrepancies in body mass index,stratification of the disease,sex,diet,and the exclusion or inclusion criteria used across studies.Significant findings in our study include an increase in the abundance ofFirmicutesandActinobacteriaand a decrease inBacteriodetesandTenericutesphylotypes in Old-HFD mice.Consistent with the obesity literature,changes in theFirmicutesphylum were positively correlated with body weight while differences in theBacteriodetesphylum were negatively associated with body weight.Since changes inFirmicutes,ActinobacteriaandBacteriodeteshave been found when comparing obese NAFLD/NASH and lean healthy children and adolescent patients,we believe that the changes observed in these phyla may be related to the body composition and not to the NAFLD pathology.At the genus level,we report an overrepresentation ofAdlercreutzia,Coprococcus,Dorea,andRuminococcusand underrepresentation ofTuricibacterandAnaeroplasmaabundance in Old-HFD mice.These findings are consistent with some,but not all,of the reported human NAFLD/NASH associated organisms.For example,Ramanet al[17]and Del Chiericoet al[65]reported elevated representation ofDoreain NAFLD/NASH children,adolescents,and adult subjects,but Da Silvaet al[63]observed lower abundance ofDoreain NAFLD patients.In obese post-menopausal women,Doreagenera has been negatively associated with insulin resistance and positively correlated with markers of inflammation[66].Taken together,these studies suggest an association between increased obesity,inflammation,and elevatedDoreagenera - which may or may not be related to NAFLD.Consistent with our findings,Boursieret al[19]and Del Chiericoet al[65]observed an increase in the abundance ofRuminococcus.Meanwhile,Da Silvaet al[63]observed that NAFLD patients had lower fecal abundance ofRuminococcus.Other genera that were altered in the Old-HFD group includingAdlercreutzia,Turicibacter,andAnaerosplasmahave not been associated with NAFLD or NASH in human patients.Thus,whether changes in abundance of these microbes is due to diet,obesity,or other factors independent of NAFLD warrants further investigation.
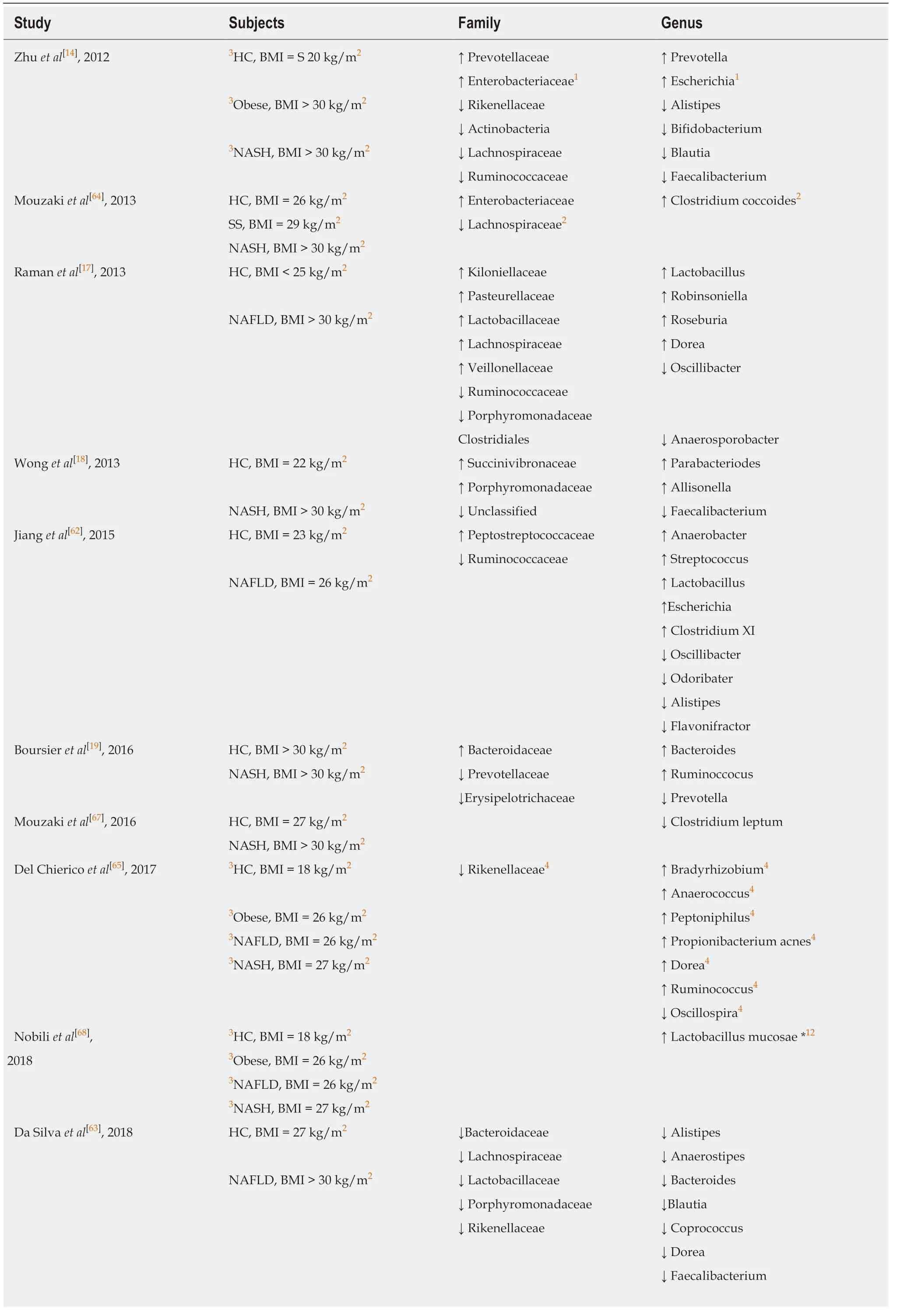
Table1 Gut microbiota associated changes in non-alcoholic fatty liver disease and Non-alcoholic steatohepatitis subjects

1Different from Obese;2Different from Steatosis or non-alcoholic fatty liver disease;3Children and adolescent;4Differences between grouping patients and healthy control.BMI:Body mass index;HC:Healthy control;NAFLD:Non-alcoholic fatty liver disease;NASH:Non-alcoholic steatohepatitis;SS:Steatosis.
A limitation in our study is that we were not able to pinpoint a causal relationship between NAFLD and specific microbes.This is in part due to an inability to dissect changes independent of long-term dietary macronutrient manipulation or obesity -both of which directly impact the composition of gut microbiota[26].In addition,it is important to note that our findings did not recapitulate all of the microbial findings that have been reported in the clinical NAFLD/NASH literature.Therefore,additional mechanistic studies are necessary to determine if any of the intestinal bacteria changes that we observed can promote NAFLD pre-clinically.Further,our study was limited to one-time point;all mice were exposed to chronic HFD feeding for the duration of 80 wk.However,it is certainly possible that a shorter duration of HFD feedings (less than 80 wk) may incite NAFLD pathophysiology;although most likely not to the same degree.It also would have been informative to examine changes across time to fully evaluate factors involved in progression of the disease.A third limitation in our study is that we are not able to conclude if the macronutrient composition of the diet is responsible for NAFLD development or rather the excess calories that intrinsically promotes NAFLD.Thus,it may be advantageous to study energy-dense isocaloric diets with high carbohydrate or fat macronutrient content.
In summary,our study examined the effects of prolonged HFD feeding on NAFLD.In particular,we focused on determining the common features of this animal model with NAFLD and human manifestation of the disease.Our prolonged HFD feeding led to the development of obesity,steatosis,non-alcoholic stetohepatitis,insulin resistance,steatosis,liver ER stress,and gut dysbiosis making it a suitable model for the study of NAFLD.Our results suggest that chronic HFD can mimic most,but not all,of the pathophysiological events observed in NAFLD.
ARTICLE HIGHLIGHTS
Research background
Animal models that can exhibit characteristics seen in non-alcoholic fatty liver disease (NAFLD)have the potential to drive the discovery of new drugs to treat this disease.
Research motivation
Most animal models used to investigate NAFLD misrepresent typical characteristics seen in human patients with NAFLD.Therefore,any successful treatments documented in these animal models may not be clinically translatable.
Research objectives
To evaluate if mice consuming a high calorie diet for a prolonged time can mimic clinical characteristics of NAFLD.
Research methods
Male mice (10 wk old) were a ssigned to the following groups:Young- low-fat diet (LFD) (n = 20;low calorie diet for),Old-LFD (n = 15;low calorie diet),and Old-HFD (n = 18;high calorie diet).Mice in the LFD consumed a diet rich in carbohydrates,meanwhile the HFD was abundant in fat content.Liver,colon,adipose tissue,and feces were collected at 16 wk of age in Young-LFD mice and at 90 wk of age in Old-LFD and Old-HFD to evaluate microscopic features,glucose metabolism,inflammation,endoplasmic reticulum (ER) stress,and microbiome profile seen in NAFLD.
Research results
Old-HFD mice showed increased body weight,blood glucose,plasma insulin,HOMA index,steatosis,hepatocellular ballooning,inflammation,fibrosis,NAFLD activity score,ER stress markers (CHOP and p-Jnk),and abundance of Firmicutes,Adlercreutzia,Turicibacter,Coprococcus,Dorea,and Ruminococcus.
Research conclusions
Mice fed a high calorie diet for 20-mo (Old-HFD) mimicked microscopic characteristic and microbial events that have been previously observed in NAFLD patients.
Research perspectives
It is important to critically select animal models to study any disease including NAFLD.Future research dedicated to investigation of new treatments for NAFLD should consider prolonged-HFD feedings as their animal model.
杂志排行

World Journal of Hepatology的其它文章
- Fascioliasis presenting as colon cancer liver metastasis on 18Ffluorodeoxyglucose positron emission tomography/computed tomography:A case report
- Outpatient telephonic transitional care after hospital discharge improves survival in cirrhotic patients
- lmpact of psychosocial comorbidities on clinical outcomes after liver transplantation:Stratification of a high-risk population
- ls porto sinusoidal vascular disease to be actively searched in patients with portal vein thrombosis?