树干横截面面积年增长量的垂直分布研究进展
2019-08-16王庆云
王庆云
(山西省林业调查规划院,山西 太原 030012)
树干横截面面积年增长量(RAI)比直径年增长量更适合表示单木及林分水平的生长量变化[1],其从梢顶到树干基部的垂直分布信息是揭示干形变化及碳分配机理,深入研究林木生长与气候关系,全面了解树干水分传输及机械支持功能的变异机制,准确预测材积年增长量和单木生物量的基础,备受树木生理学、森林生态学、木材学等学科学者的关注[2,3]。笔者就RAI垂直分布研究的意义、现状和方向等进行了综述,以期为全面了解该方面的研究进展提供参考。
1 RAI垂直分布的观测方法
树干横截面面积年增长量(RAI)垂直分布的观测方法主要包括2种,一种是在不同树干高度处逐年实测RAI,如,Sumida[4]等对日本扁柏的树干基部、0.3 m树高处、其余部位每隔1.0 m的RAI进行了连续20 a的测定,研究了其垂直分布的年际变化;另一种是在不同树干高度处截取圆盘或钻取样芯,采用年轮分析仪等测算各位置、逐年的RAI[5],但不同研究选用的采样位置和间隔等差别较大(第32页表1)。此外,在第2种方法中,不同研究选用的RAI测算时长也不同,有的以单个年份,有的以最近3 a[6],5 a[7],甚至36 a[3]的年均增长量作为RAI.可见,目前尚无统一、规范的RAI采样和测算方法,从而导致不同的研究结果难以比较,由此造成的影响尚不明确[5]。
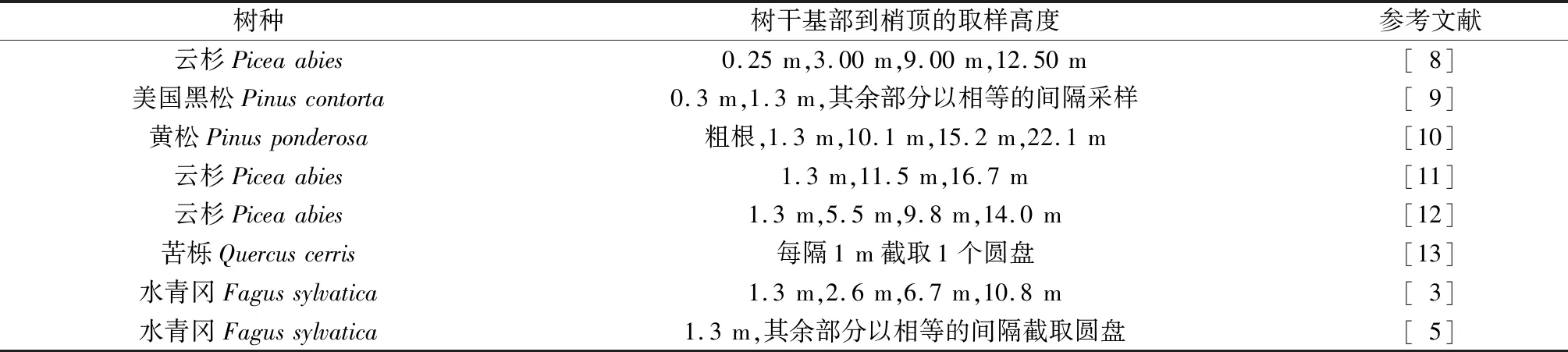
表1 不同研究的采样方法
2 RAI垂直分布的理论和实际模式
2.1 RAI垂直分布的理论模式
树干的主要功能是水分传输和机械支持。一些研究根据相关理论提出了相应的RAI垂直分布模式,即理论模式。目前,RAI垂直分布存在多种理论模式,这些模式间尚存分歧。管道模型理论将树干概化为管道集合体,认为单位输水管道支持单位叶面积[14]。阔叶树种树干的有效输水组织为其木质部最外侧的1个年轮,针叶树种为其最外侧的几个年轮层,其数量与针叶寿命一致[15,16]。据管道模型理论,树干某位置RAI与其上方的当年生叶量或总叶量成正比[15]。该模式已成为单木生长模型中确定和模拟垂直分布的主要依据[17]。有研究认为,根据管道模型理论推断,RAI垂直分布并未考虑水分传输阻力的影响[7]。据水力限制及其补偿机制假说[18-20],水分传输阻力与传输长度成正比,与传输组织直径的4次方成反比,林木通过改变传输组织直径等,使传输阻力随树高增加而下降[19],以保证所有叶片的水分供应[21]。据此推断,RAI随树高增加而减小。WBE最优水分传输系统理论认为,水分传输组织随分枝级数的增加而均匀细化[22,23],据此推断,冠下RAI恒定,冠内RAI以节间(相邻轮生枝间的树干)为单元,随树高增加而均匀细化。均匀应力理论认为,为保持树干直立,保证树体在风力等作用下能恢复到正常位置,提高其对弯曲荷载的抵抗力,防止应力垂直分布不匀导致树干断裂等,树木会根据应力的垂直分布调整RAI的垂直分布,以保证应力分布的均匀性[24]。Dean等[25]基于均匀应力假说,构建了不同树干位置的RAI模型。据此模型,RAI随树高增加而减小。
2.2 RAI垂直分布的实际模式
实测结果表明,RAI从梢顶到树干基部的垂直分布可分为3个区段,依次为有效树冠区、中间区和膨大区。其中,有效树冠区是指林分郁闭后,生长和光合作用相对旺盛的林木中上部树冠,受株间竞争等影响较小;而下部由于受到遮阴和挤压成为衰退树冠[26]。一般通过树干解析和RAI分段拟合的方法可以确定有效树冠长度[27]。RAI垂直分布模式的株间差异主要源于树干中间区。据此差异,RAI垂直分布的实际模式主要包括3种。第1种为RAI在有效树冠区、中间区和膨大区分别以相对中等、较小和较大的斜率随树高增加而减小;第2种为RAI在有效树冠区、膨大区分别以相对中等和较大的斜率随树高增加而减小,在中间区保持恒定;第3种为RAI在有效树冠区、膨大区分别以相对中等和较大的斜率随树高增加而减小,在中间区以相对较小的斜率随树高增加而增加。其中,膨大区相对高度一般为5.0%[28]~10.0%[6],绝对高度随胸径增加而增加。
2.3 理论和实际模式的差别
Gaffrey[7]对RAI垂直分布的理论和实际模式进行了比较,发现任何一种理论模式均不能完整解释RAI的实际垂直分布模式,只能解释部分树干区段的变异。如,管道模型理论只能解释冠内RAI的垂直分布,均匀应力理论只能解释膨大区的垂直分布。
3 RAI垂直分布模拟方法
Courbet[26]研建了一个灵活的3分段模型,允许RAI在冠内非线性变化,在树干中间区线性变化,在树干基部剧增,并提供了直观、无相关性的模型系数,且允许在不同生长条件下对系数进行调整。该模型适用于单木尺度,但叶量等输入变量不易获取。此外,一些研究基于管道模型理论构建了的RAI和叶面积的关系模型[17],基于光合产物分配的半机理模型[17,27,29],描述了光合产物在不同树干高度的收支、分布和转移,然而不能模拟预测膨大区RAI的垂直分布。Cortini[6]等在总结这些模型优缺点的基础上,以易于获取的因子为自变量构建了一个3分量模型,在加拿大4个针叶树种上的应用取得了理想效果,并认为该模型的建模方法和形式适用于多数树种,但实际效果尚需验证。
4 RAI垂直分布的主要影响因子
除水分传输和机械支持功能外,遗传、生理生态因子、森林培育和经营措施等对RAI垂直分布的影响也不可忽视。耐阴树种和阳性树种的RAI垂直分布模式不尽相同,主要是因为树种的耐阴性影响活树冠内叶的垂直分布,在相同的竞争条件下,耐阴物种的冠幅更大[30,31]。在生长较快的树木上,形成层生长素浓度垂直变化明显,沿树高显著增加;但在生长较慢的树木上却无变化,继而影响不同林木的RAI垂直分布[32,33]。在瑞典北部,对45年生樟子松进行施肥和间伐处理,结果发现,间伐1 a后,径向生长量增加,然而施肥后的第2年径向增长量才开始增加,这是由于间伐主要促进下部树干生长,而施肥主要促进上部树干生长[34]。林木竞争和优势度影响RAI垂直分布,如,优势木、中间木和被压木RAI的垂直分布分别为第一、第二和第三种分布模式[4]。不同树干位置RAI对气候变化的响应不同,胸高处的敏感性高于其它位置[9]。此外,冠基抬升速度和过程也影响RAI的垂直分布[4,35,36]。这些研究成果为解释RAI垂直分布模式的成因奠定了良好基础,但这些因子的单独和综合影响目前尚不完全清楚[17]。
5 与代表性观测位置的关系
在树木径向生长的常规测量及树木年代学研究中,普遍在胸高处测定径向生长量或取样(钻取样芯或截取圆盘),其基本假设是该处的RAI能代表全树干水平的径向生长量,或至少能代表全树干或单木水平径向生长对气候变化的响应[37]。所谓代表性观测位置就是该位置的RAI和全树干水平的RAI最为接近,全树干水平的RAI通常以树干不同位置平均或加权平均的RAI表示。对垂直分布模式的年际变化研究发现,在正常年份,胸高处与全树干水平的生长变化趋势一致;但在气候胁迫年份,如,干旱年份,垂直分布模式发生变化,胸高处的代表性明显下降,此时以该处观测结果推断全树干水平的RAI会造成较大误差[5]。此外,随着林木的生长发育,RAI的垂直分布模式及代表性观测位置也不断变化,如,欧洲山毛榉(Fagussylvatica)在6.7 m处[3],北美红杉(Sequoiasempervirens)在15.0 m处。因此,基于RAI垂直分布信息,正确判断代表性观测位置及其变异规律,对提高全树干水平RAI估算的准确性十分关键。
6 RAI与VAI和单木叶生物量的关系
不同树干位置RAI与全树干材积年增长量(VAI)及单木叶生物量间的关系备受关注。绝大多数研究认为胸高处的RAI与VAI关系紧密,二者成显著、稳定的线性函数关系[13]。但有研究发现二者关系并不确定,有时以乘幂函数描述更为合适。还有研究发现,受垂直分布模式的影响,树冠基部的RAI与VAI关系更为紧密[5]。因此,在构建RAI和VAI关系模型时,RAI垂直分布模式对理想位置及二者关系的影响不容忽视。
RAI垂直分布模式不同,各位置RAI与叶生物量关系的一致性存在差异[22]。多数研究认为活树冠基部比其它位置的边材面积对单木生物量的预测效果要好[38,39]。但有研究发现各模式及整体水平,胸高处均是预测叶生物量的理想位置,活树冠基部仅在树干中间区RAI较为恒定的模式中具有较好的预测效果[40]。
7 研究展望
从RAI垂直分布的研究进展可知,目前其株内和株间变异机理尚不完全明确,确定合理的采样和观测方法,将水力学、生物力学及生理学过程相结合,阐明关键影响因子的作用机制是揭示其内在机理的关键[16]。
RAI的垂直分布信息是确定RAI的代表性观测位置、基于RAI预测VAI(单木材积增长量)和单木叶生物量的理想位置的重要依据,但分布模式对这些位置和相关模型的影响尚不完全清楚,对此开展研究具有重要的理论和实践价值,是未来研究的重点。
