古槐树叶片主要性状多样性分析
2019-08-08楚晓晓李双云周继磊刘盛芳庞彩红
楚晓晓,杨 勇,李双云,夏 阳,周继磊,刘盛芳,屈 星,庞彩红
(1.山东省林业科学研究院,山东 济南 250014;2.山东省林木遗传改良重点实验室,山东 济南 250014;3.甘肃农业大学园艺学院,甘肃 兰州 730000;4.平凉职业技术学院,甘肃 平凉 744000;5.山东省林木种苗和花卉站,山东 济南 250014)
槐Sophora japonica为豆科Leguminosae槐属Sophora落叶乔木树种。原产中国,现南北各省区广泛栽培,华北和黄土高原尤为多见,对土壤要求不严,适应能力强,在石灰性、酸性及轻盐碱土上,甚至在山区水少的地方均可正常生长。此外槐冠大荫浓,具有抗污染,耐烟尘,抗风等特点,是良好的遮荫树和行道树,槐的花、枝、果实均可入药,具有良好的开发利用前景[1-2]。
山东省古槐树的数量居所有古树名木之首[3],绝对数量多,古树树龄长(300~1 800a),分布范围较广,加之当时的交通不发达,使古槐树的基因型多样性成为可能。另外,古树历经数百年乃至千年自然环境的考验存活下来,表明其对逆境具有良好的抵抗能力,因此开展古槐树种质资源的调查、收集、多样性分析具有重要的现实意义,也为开展槐逆境胁迫适应机制、高抗种质材料早期诊断评价与优选技术研究提供材料,为槐杂交育种和分子育种奠定基础,为新品种的选育提供种质背景。
近年来对槐研究多集中在催芽[4]、扦插[5-6]、嫁接[7-8],组培[9]等方面,对叶表型多样性分析还未见报道。叶片是高等植物进行光合、呼吸及蒸腾作用的主要器官,而叶外部形态特征与该植物的生理营养及其所在的生态地理因子有关[10],其表型的变化将直接影响植物体的一切生理活动,因此叶表型性状多样性是遗传物质多样性的具体表现,是进行遗传多样性研究最基本、最重要的组成部分之一[11]。本试验对72份古槐树种质叶表型性状遗传多样性进行分析研究,为槐的种质资源收集保存、鉴定分类、良种选育、栽培研究及开发利用提供参考依据。
1 材料和方法
1.1 供试材料
72份古槐树种质(树龄均大于300年),来源于临沂(LY)、烟台(YT)、枣庄(ZZ)、淄博(ZB)四个地级市的不同县、市、区,每个地级市18个无性系,2011年春天收集古树上长势好,无虫无病的枝条,于当年四月底五月初进行嫁接,选取带有2~3个饱满芽的接穗,嫁接于1年生的实生苗砧木上,每个无性系嫁接10株,嫁接成活植株定植于山东文峰集团(威海市)苗圃(121°98′E,37°31′N),2012年定植。
1.2 实验方法
1.2.1 样品采集2016年8月,每份种质选取正常生长的植株3株,每株外围取4个成熟枝(不同方向),从成熟枝条中部选取发育正常的一枝复叶,共计12个复叶,用于小叶数的统计和叶轴长的测定;选取复叶上(从顶叶数)第三对小叶,每份种质共计24片叶片用于小叶叶片性状的观察与数据的测定。
1.2.2 测定性状小叶叶片的干质量(X1)、鲜质量(X2)、叶脉角(X3)、叶周长(X4)、叶面积(X5)、叶厚(X7)、叶长(X8)、叶宽(X9)、叶形指数(X10)、小叶数(X11)共10项,此外还有复叶叶轴长(X6),共计11个性状。
1.2.3 测量方法叶厚测定 用游标卡尺测量叶片厚度,6片一组(24片叶片随机分成4组,每组6片),测量后取平均值(精确到0.01mm)[12];鲜质量测定:样品采集后立即用METTLER TOLEDO(JE703CE)电子天平称其鲜质量(精确到0.01g);干质量测定:将叶片置于烘箱中烘至恒质量,METTLER TOLEDO(JE703CE)电子天平称其干质量(精确到0.01g);叶轴长测定:利用直尺测量叶轴的长度(精确到0.01 cm);小叶数测定:通过目测进行观测(片);叶形指数测定:是叶长与叶宽的比值[13]。叶长、叶宽、叶周长、叶面积、叶脉角测定:利用EPSON V370扫描仪,获取图像,将待测叶片平铺到扫描仪面板上,盖上玻璃盖板,使叶片完全平整展开,叶正面朝下;图像类型为48位全彩,分辨率为300 dpi,文稿大小宽度215.9 mm,高度297.2 mm,图像格式为bmp格式[14]。Digmizer4.2图像分析软件对扫描图像进行分析,以图片高度297.2 mm设置比例尺,测量叶面积(0.01 cm2)、叶周长、叶长、叶宽精确到(0.01 cm);叶脉角利用软件上的角度测量其叶脉角。
1.3 数据分析
利用Excel对原始数据的平均值、最大值、最小值、标准差、变异系数等进行整理统计;用SPASS软件进行方差分析、主成成分分析、相关性分析和聚类分析。
2 结果与分析
2.1 古槐树叶片性状的多样性
数量性状可用来分析遗传变异的水平[15],一般认为变异系数大于10%就说明样本间的差异较大,变异系数越大,个体之间的差异也就越大,越有利于对种质资源进行鉴定、评价和利用[14]。对72份古槐树叶片的11个性状指标进行了统计分析,结果如表1。由表1可知,古槐树叶片的干质量、鲜质量、叶脉角、叶周长、叶面积、叶轴长、叶厚、叶长、叶宽、叶形指数、小叶数的平均值分别为0.10 g,0.28 g,53.15°,16.40 cm,10.95 cm2,15.31 cm,0.28 mm,6.06 cm,2.49 cm,2.47,14片。变异系数11.88%~39.10%,平均变异系数23.26%。在所测种质叶片性状中,鲜质量最大为0.77 g,最小为0.05 g,变异范围最大,变异系数高达39.10%;叶厚的变异系数最低,11.88%,古槐树的叶片性状变异系数从小到大依次为:叶厚<叶脉角<叶形指数<叶轴长<小叶数<叶宽<叶长<叶周长<叶面积<叶干质量<叶鲜质量。经过F值检验,11个叶片性状均达到了极显著差异(P<0.01),表明古槐树叶片表型数量性状变异范围较大,差异显著,遗传变异水平较高,遗传多样性较丰富[16-17]。

表1 古槐树叶表型性状变异分析Table1 Analysisonphenotypic trait variation of old S.japonicaleaves
2.2 古槐树叶片不同性状间的相关性
相关性分析是指对两个或多个具备相关性的变量元素进行分析,衡量两个变量因素的相关密切程度。对72份古槐树种质叶片性状进行相关性分析,结果如表2。

表2 叶片不同性状间的相关性分析Table2 Correlation analysis ondifferent leaf traits
由表2可知,叶面积与干质量、鲜质量、叶周长的相关系数达到了显著性正相关,其相关系数分别为0.63,0.83,0.68。叶宽与干质量、鲜质量、叶脉角、叶周长、叶面积、叶长达到了极显著(P<0.01)和显著(P<0.05)正相关,其相关系数分别为0.56,0.72,0.10,0.58,0.88,0.61。叶长与干质量、鲜质量、叶周长、叶面积存在显著或极显著正相关,其相关系数为0.51,0.67,0.83,0.77,叶周长与小叶数、鲜质量呈极显著正相关,相关系数为0.43,0.58。叶形指数与叶周长、叶长之间存在极显著正相关,其相关系数为0.30,0.48。小叶数与叶轴长之间的相关和干质量与鲜质量之间的相关存在极显著正相关,其相关系数分别为0.63和0.60。另外,叶宽与叶面积相关系数达到极显著,其相关系数最高0.88。结果显示,不同性状间存在一定的相关性,相关系数表明基于一个性状的选择也可能同时改良另一个性状[18],因此,在筛选与叶面积和叶周长相关的品种时,可以参考叶长和叶宽指标。在对多个叶片性状进行改良时,应关注性状间的相关性并充分利用这些相关性来提高育种效率。
2.3 古槐树叶片性状的主成分分析
通过主成分分析将11个性状进行线性组合,构成少数不相关的新变量,原变量中的信息被新变量代替。将原始数据矩阵经过标准化后,进行主成分分析,通过分析,能更清楚地显示各性状在叶片表型性状多样性构成中的作用。
通过分析提取了4个主成分(表3),其累计贡献率达到75.70%,包含了11个指标的大部分信息,因此选取前4个主成分作为槐叶片性状选择的综合指标。在第1主成分中叶面积(PC1)占最高系数,方差贡献率为37.85%,说明第1主成分是表示叶面积性状的综合因子;在第2主成分中叶轴长(PC2)占有最高系数,方差贡献率为15%,说明第2主成分是表示叶轴长性状的综合因子;在第3主成分中叶形指数(PC3)占有最高系数,方差贡献率为12.97%,说明第3主成分是表示叶形指数性状的综合因子;在第4主成分干重(PC4)占有最高系数,方差贡献率为9.71%,说明第4主成分是表示干重的综合因子。由此可知,槐叶片之间的差异主要表现为叶面积、叶轴长、叶形指数、干重的不同。这些主成分在槐种质分类上起到了重要作用,这些为槐新品种的选育过程中确定育种目标,提供了依据。
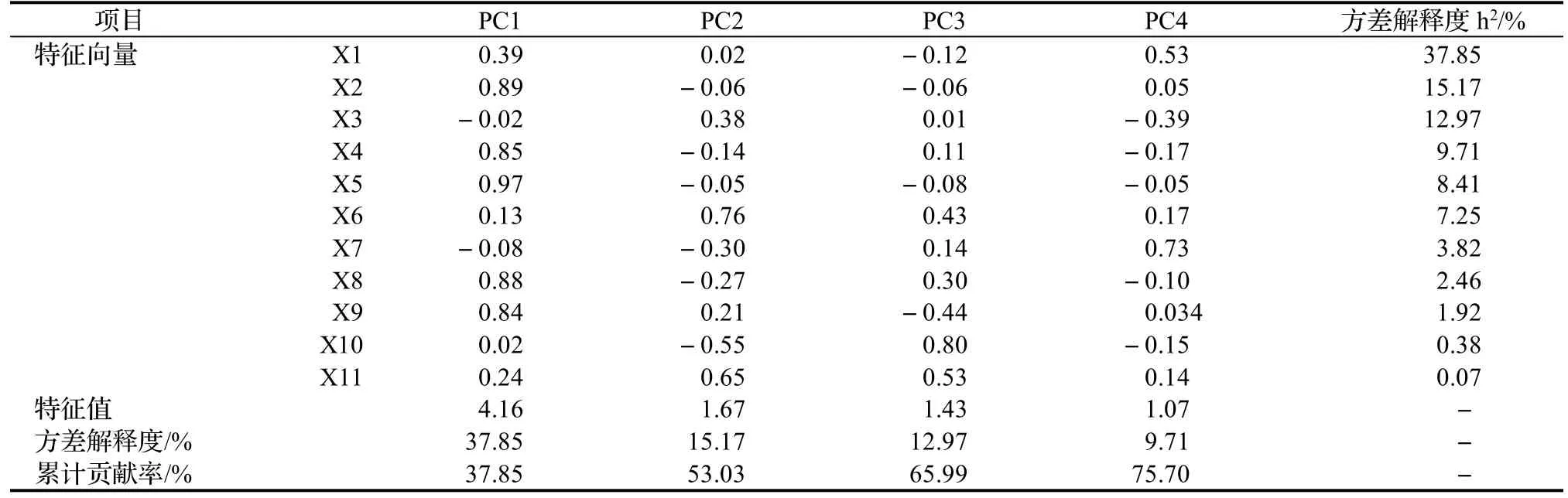
表3 叶片性状主成分分析Table3 Principal componentanalysisonleaf phenotypic traits
2.4 古槐树综合性状的聚类分析
对72份古槐树种质叶表型的11个性状的原始数据进行标准化,并计算欧式距离,利用可变类平均连接法(组间)对其进行聚类分析,将72份种质完全区分开,说明叶片性状能够反映各样品间的差异(图1),来自同一种源的种质没有完全聚到一起,而是相互交叉。
由图1表明,取阈值在22.5时,可分为3大类群,分别以Ⅰ、Ⅱ、Ⅲ命名(表4)。三大类群中,第Ⅰ类群包含2份种质,约占总样本数的2.8%,与其它两类群相比较,叶周长、叶长的平均值最大,分别为21.09 cm,6.62 cm。第Ⅱ类群包含4份种质,约占总样本数的5.6%,与其它两类群相比较,叶轴长、叶宽、叶形指数、小叶数的平均值最大,为18.6 cm,2.82 cm,2.87,13片。其余66份材料的为第Ⅲ类群,约占总样本数的91.6%,鲜质量、干质量、叶脉角、叶面积的平均值最大,分别为0.29 g,0.10 g,53.59°,11.09 cm2。由此可见,72份古槐树种质资源具有各自的叶片表型特征。

图1 叶片主要性状聚类图Figure1 Dendrogram of leaf traits of S.japonica

表4 各类群的平均值Table4 Mean phenotypic traits of different groups
3 结论与讨论
种质资源遗传多样性的研究能够为亲本选择、杂交组合配制以及遗传改良提供有效的信息。形态学方法是植物遗传多样性研究最直接、基础的方法,表型多样性是在形态水平上对遗传多样性进行的阐述[10,17,19-20],很多学者先后利用表型性状对种质资源的多样性进行了分析[21-24]。如万继峰等[25]利用了12个叶片表型性状进行了橄榄Canarium album种质表型多样性研究,叶片表型变异系数为9.52%~25.09% ;徐斌等[18]利用14个叶片表型性状对杜鹃红山茶Camellia azalea的多样性进行分析,叶片表型变异系数为5.30%~47.00%,变异系数差值比较大。本次研究了72份古槐树种质11个叶片表型性状的多样性,其变异系数为11.88%~39.10%,且在0.01水平上达到了极显著差异,结果说明叶片存在着丰富的多样性,不同植物叶片的变异程度大小不同。宋伟栓[26]利用SRAP标记分析了32份不同来源地的古槐树遗传距离和相似系数,遗传距离在0.104 0~0.916 3之间,平均值为0.397 9,遗传相似系数变化范围为0.333 3~0.888 9,平均值为0.619 0,说明古槐树间存在一定程度的遗传变异,但遗传变异程度不是很大,这与叶片表型变异程度不太一致,可能与样本量有一定关系,也可能是外部环境对表型的影响造成的。
基于表型性状对古槐树种质资源进行聚类分析,在欧式距离为22.5时,72份槐种质资源分为3大类群,发现同一地理种源的材料并未聚到一起,这与宋伟栓[26]的研究结果是一致的,说明聚类结果与各种源的地理位置分布不相符,即叶片性状与种源无直接关系,出现这一结果的原因可能与气候、土壤、海拔、以及引种等因子对表型的影响有关,有待进行进一步研究。
古槐树的数量居山东省所有古树名木之首,存在时间长,分布范围较广,基因型众多,是具有创新利用潜力的种质资源,由于古槐树特定的文化底蕴使得大量的古树资源被保存下来,扩大了种质基因库;另一方面对古槐树表型性状多样性的分析,揭示了各个性状之间的相关性,明确了各成分的贡献率,为古槐树表型新品种筛选与培育提供了参考,对于单株显著特异性的,可进一步研究挖掘,探寻潜在开发利用价值,同时,根据育种目标选择性状互补的亲本杂交组合,加速槐遗传育种工作进程,为种质资源的保护、利用和创新提供物质基础,也为古槐树保护重点的划定提供参考。
