向日葵MADS-Box基因HAM23-like克隆和表达分析
2019-08-08苏周吴雨雷豆韦小英何卓远杨军邹建
苏周,吴雨,雷豆,韦小英,何卓远,杨军,邹建
(西华师范大学西南野生动植物资源保护省部共建(教育部)重点实验室,四川 南充 637009)
花器官发育是陆生植物在生殖进程中的重要过程,该过程对于开花植物的有性生殖具有极其重要的意义。在进化过程中,由于自然选择的作用和花器官的起源不同,不同开花植物之间形态上存在多样性[1-3]。典型的双子叶植物的花从外到内由4轮不同的花器官组成,第1轮是萼片,第2轮是花瓣,第3轮是雄蕊,第4轮是心皮[4]。经典的ABC模型将假定参与花发育的基因分为ABC 3类。其中,A类基因控制萼片和花瓣,该类基因功能缺失将导致萼片和花瓣变成心皮和雄蕊;B类基因控制花瓣和雄蕊,该类基因失活花瓣和雄蕊将变成萼片和心皮;C类基因控制雄蕊和心皮,该类基因的缺失将引起雄蕊和心皮变成花瓣和萼片[4]。现有的研究显示,ABC模型中参与花器官发育调控基因中大多数都属于MADS-box基因家族[5]。
MADS-box基因家族在动物、植物和真菌中广泛存在,其在植物生长发育的各阶段发挥着重要的调节作用[6-7]。MADS 是MCM1、AGAMOUS、DEFICIENS和SRFS的首字母缩写组成。其中MCM1(minichromosome maintenance1)的功能是调节细胞周期、生长、代谢、专一性和决定细胞类型[8];AGAMOUS(AG)和DEFICIENS(DEF)均是花器官基因的编码蛋白,分别来自拟南芥(Arabidopsis thaliana)和金鱼草(Antirrhinum majus);SRF(serum response factor)为人类血清应答因子[9-11]。这些基因均具有一个由56~58个氨基酸组成的高度保守结构域,称为MADS-box结构域,故将其命名为MADS-box基因[12-13]。MADS-box基因不仅参与调控开花时间,决定花分生组织的发生和花器官特征,而且在根、叶、胚珠和果实的发育过程中也起着重要作用[6-13]。Koo等的研究结果显示,过表达AGL6基因会造成拟南芥提前开花[14];此外番茄中的研究证明了MADS-box家族成员rin基因参与果实成熟的调控,而SlMBP21基因则控制番茄萼片的大小[15-16]。
向日葵(Helianthus annuus)是典型的菊科(Compositae)植物,头状花序由外轮不育的舌状花(缺乏雄蕊和雌蕊)和内部可育的管状花组成。迄今为止,在形态学方面对菊科植物花形态和发育进行了大量研究,但从分子遗传学的角度来看,这一种类极多的被子植物科的花序和花的发育仍不清楚,而MADS-box基因在影响花形成的所有阶段都发挥着重要的作用[17]。到目前为止,关于向日葵的MADS-box基因研究较少,在控制花序形态发生方面,仅有9个向日葵MADS-box基因被分离和报道,包括2个A类基因HAM 75和HAM 92,影响花瓣和种皮的形成;4个B类基因HAM31、HAM2、HAM63和HAM91;2个C类基因HAM45和HAM59,HAM59参与花分生组织的终止,并与HAM45共同确定雄蕊和雌蕊;1个E类基因HAM137[17-18]。研究向日葵的MADS-box基因,能深入了解MADS-box基因对向日葵营养生长和生殖生长的影响。目前对向日葵的MADS-box transcription factor 23-like基因尚未见研究报道。本文从向日葵花中克隆了1个与花发育相关含有MADS-box结构域的基因,通过NCBI对其进行序列比对,确定为向日葵MADS-box transcription factor 23-like(HAM23-like),利用在线生物软件对其编码蛋白进行理化性质和生物信息学分析,随后采用实时荧光定量PCR(qRT-PCR)对该基因的表达模式进行分析,为探究HAM23-like基因的功能及MADS-box基因对向日葵花发育的调控作用奠定基础。
1 材料和方法
1.1 材料
试验材料是向日葵光雾山野生葵品种(Helianthus annuus‘GWMountainWild’),种于温室(光 /暗 =16 h/8 h,28℃/24℃)中。分别采集根、茎、叶、花、果实(MSt,籽粒饱满阶段)后用液氮速冻,置于-80℃冰箱保存。
E.Z.N.A®Plant RNA Kit购于OMEGA;荧光定量染料试剂SYBR®Premix ExTaqTMⅡ、反转录试剂盒PrimeScriptTMRT Reagent Kit With gDNA Eraser(Perfect Real Time)等购自宝生物(TaKaRa)公司;FastPfuFly DNA聚合酶、克隆载体pEASY®-Blunt Cloning Kit、大肠杆菌DH5α购于北京全式金。其余化学试剂均为国产分析纯级。
1.2 总RNA提取和合成cDNA
在开花前 35(-35)、-25、-15、-10、-5、0 d(开花当天)、开花后5 d(未成熟果实)采集花;并在向日葵开花当天分别采集根、茎、叶、果实(籽粒饱满阶段)及6种花器官:苞片、雌蕊、冠毛、花瓣、雄蕊、子房。材料采集后,按照E.Z.N.A®Plant RNAKit试剂盒说明书提取各组织的总RNA,然后用琼脂糖凝胶电泳和NANODROP 2000c检测提取RNA的浓度和纯度。以所得RNA为模板参照TaKaRa反转录试剂盒说明书进行反转录,合成cDNA。
1.3 基因克隆
根据向日葵全发育时期表达谱库454和向日葵花发育时期转录组数据库(均未上传NCBI)筛选到的基因序列,并在NCBI(https://www.ncbi.nlm.nih.gov/)中比对获得目的基因登录号(XM_02213319 8.1),根据基因序列设计特异性引物transcription factor 23-F/R,以反转录第一链cDNA为模板,扩增transcription factor 23基因全长序列。PCR扩增体系共 50μL,包含 cDNA 模板 3μL,transcription factor 23-F/R 引物各 1μL,5×buffer 10μL,dNTP 4μL,ddH2O 30μL 和 FastPfu Fly DNA 聚合酶 1μL,均加入200μL离心管中,用枪尖冲打混匀。扩增程序为98℃ 30 s,56℃ 30 s,72℃ 1.5 min,34 次循环,72℃延伸5 min。扩增产物经1%的琼脂糖凝胶电泳检测,切胶回收,连接到pEASY®-Blunt vector克隆载体,并转入大肠杆菌(Escherichia coli)DH5α感受态细胞中,在含100 mg L-1Kan的LB培养基中37℃恒温培养过夜,筛选出阳性转化细胞,送至上海生物工程公司测序。
1.4 系统发育树的构建
利用NCBI中在线软件ORF Finder对所有拟南芥和向日葵MADS-box基因序列进行分析,推导出编码的氨基酸序列。拟南芥MADS-box基因由拟南芥数据库获得利用MEGA 6.0软件构建拟南芥(https://www.arabidopsis.org/)和向日葵 MADS-box的系统发育树,预测向日葵HAM23-like基因与拟南芥的亲缘关系。
1.5 HAM23-like的生物信息学分析
利用在线软件ProtScale(http://web.expasy.org/protscale)和 ExPASy(http://web.expasy.org/protparam/)分别对HAM23-like基因编码蛋白的亲/疏水性和理化性质进行预测。
使用DNAMAN 8.0软件对HAM23-like、拟南芥Agamous-like 18的氨基酸序列和朝鲜蓟(Cynara cardunculusvar.scolymus)、莴苣(Lactuca sativa)和番茄(Lycopersicon esculentum)等高等植物的MADS-box transcription factor 23-like基因编码的氨基酸序列进行同源比对。
1.6 HAM23-like的表达模式
采用实时荧光定量法检测,利用Primer primer 5.0设计HAM23-like基因的实时荧光定量引物,以向日葵eF1A为内参基因(表1)。以提取的向日葵cDNA为模板进行qRT-PCR扩增,扩增体系为:SYBR®Premix ExTaqTMⅡ 5μL,Transcripition factor 23-F/R各 1μL,ddH2O 2μL,模板 cDNA 1μL。PCR 反应在CFX96 Real-Time PCR仪(Bio-Rad,美国)上完成,3次重复。先确定最适解链温度(melting temperature,Tm),再构建标准曲线,最后进行检测。
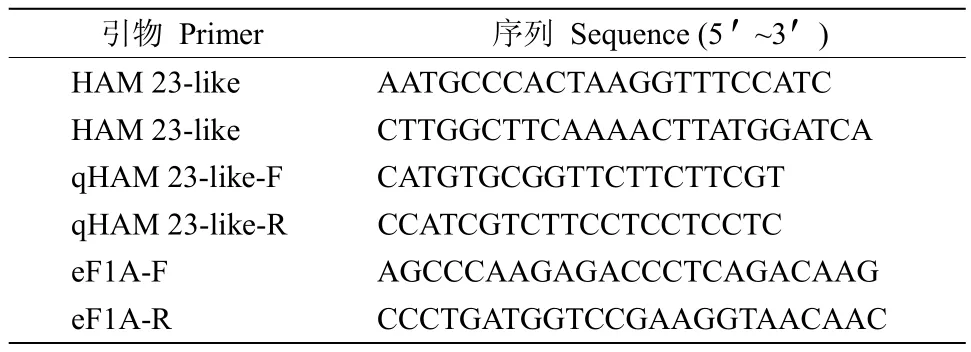
表1 引物序列Table1 Primer sequence
2 结果和分析
2.1 HAM23-like的克隆
以向日葵总RNA反转录所得cDNA为模板,用HAM23-likeF/R引物进行PCR扩增,得到与预测大小一致的条带(图1)。为了验证克隆准确性,挑选10个单克隆菌落进行测序,测序片段为956 bp,包含了HAM23-like基因完整的开放阅读框(ORF)。
2.2 HAM23-like的生物信息学分析
从NCBI中BLAST比对找到HAM23-like基因的序列,再通过NCBI的CD Search搜索HAM23-like基因编码蛋白的保守结构域,结果表明HAM23-like的ORF为831 bp(图2),编码276个氨基酸。该基因编码的蛋白具有保守的MADS-box和K-box结构域,分别位于氨基酸序列的第46~119和128~211位(图3)。利用在线分析网站ExPASy对HAM23-like蛋白进行组分分析,相对分子质量为30.52 kD,理论等电点为9.42,属于不稳定蛋白,不稳定指数为55.03;脂溶性指数为76.03;亲水性平均值(GRAVY)为-0.683。蛋白中包括正电荷氨基酸残基(Arg+Lys)46个,负电荷氨基酸残基(Asp+Glu)35个,分别占总氨基酸数的16.7%和12.7%。疏水性最强和亲水性最强的氨基酸分别是位于第91位的Ile(1.944)和264 位的 His(-3.778)(图 4)。

图1 HAM23-like的扩增。M:DL2000 DNA Marker。Fig.1 PCR of HAM23-like.MDL2000 DNAMarker.
将推测的向日葵HAM23-like基因与其他植物MADS-box基因编码的氨基酸序列进行同源比对,结果表明HAM23-like具有典型MADS-box保守结构域,即MIKC结构域。随后构建了基于MADS-box基因编码氨基酸序列的拟南芥和向日葵系统发育树(图5),包括9个已报道的向日葵MADS-box氨基酸序列和本研究挑选出的8个疑似与花发育相关的MADS-box氨基酸序列。结果表明,HAM23-like蛋白与拟南芥的AGL18蛋白聚在同一分支上,说明两者有较高的亲缘关系。
将HAM23-like基因与拟南芥Agamous-like 18、朝鲜蓟、莴苣和番茄的MADS-box transcription factor 23-like基因等编码的氨基酸序列进行同源比对,结果表明(图6),HAM23-like在第46~119位有典型的MADS-box保守结构域,拟南芥AGL18蛋白和朝鲜蓟、莴苣及番茄的MADS-box transcription factor 23-like蛋白的保守结构域分别位于第2~75、2~74、43~115和 2~77位,说明 MADS-box蛋白的保守结构域具有高度相似性。同时,HAM23-like与朝鲜蓟的MADS-box transcription factor 23-like相似度最高,达到81%;可能向日葵与拟南芥属于不同科,存在一定的差异,与拟南芥的AGL18的相似度不高。
2.3 HAM23-like的表达分析

图2 HAM23-like基因序列和编码的氨基酸序列Fig.2 Sequence of HAM23-like and encoded amino acid sequence

图3 HAM23-like蛋白的保守结构域Fig.3 Conversed domain of HAM23-like protein
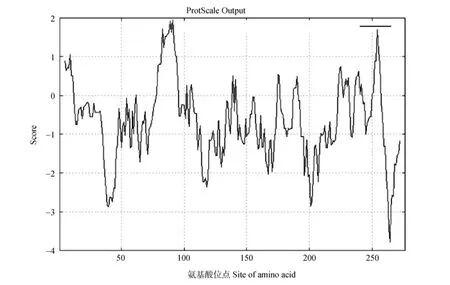
图4 HAM23-like蛋白的亲、疏水性预测Fig.4 Prediction of hydrophobicity and hydrophilicity of HAM23-like protein

图5 拟南芥和向日葵MADS-box蛋白的系统发育树。A:拟南芥;c:向日葵。Fig.5 Phylogenetic tree of MADS-box proteins in Arabidopsis and Helianthus.A:Arabidopsis thaliana;c:Helianthus annuus.

图6 向日葵HAM23-like的氨基酸序列与其他植物的同源性比较Fig.6 Homology comparison of Helianthus annuus HAM23-like protein with other plants
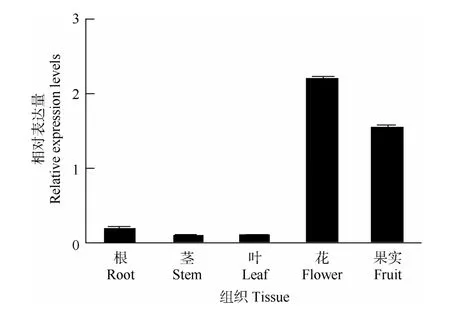
图7 HAM23-like在不同组织中的表达Fig.7 Expression of HAM23-like in different tissues
组织表达模式分析结果表明(图7),HAM23-like基因在向日葵的根、茎、叶、花和果实中均有表达,在花的表达量最高,其次是果实,茎中的表达量最低。HAM23-like基因在花中的最高表达量达到了茎中表达量的24倍。这说明HAM23-like可能在花发育和果实的形成过程中起重要的作用。
对花发育6个时期(-35至0 d)和果实发育早期(5 d,未成熟果实)及开花当天的6个花器官中HAM23-like基因的表达量进行分析,结果表明(图8),花发育的整个过程中,HAM23-like基因的表达量逐渐升高,尤其在开放前5 d(-5 d)表达量激增,并在完全开放时达花期最高值,在果实发育早期HAM23-like基因表达达到整个生命周期的峰值,这表明HAM23-like可能在向日葵开花过程和果实发育早期发挥重要的调控作用。同时,在开花当天,HAM 23-like在雄蕊和子房有表达,且雄蕊中的表达量极高,这说明HAM 23-like可能对雄蕊和花粉的形成起重要的调控作用,也对果实早期的形成有着一定的影响。
3 讨论
高等植物生活周期史中,生殖发育或花发育是极其重要的一个环节,受到遗传因素和外界环境的双重影响,是开花基因在时间和空间顺序上的表达与外部环境的影响共同作用的结果[19-20]。植物花发育进程涉及到光周期、春化、自主和赤霉素等信号途径[21]。此外,MADS-box家族转录因子的调控作用也是该进程不可或缺的因素[22]。然而,MADS-box家族对植物生长的调控作用体现在多个方面,不仅参与花分生组织的发生和花器官形态建成的调节[23],还与控制开花时间[24-31]、调节侧根生长[32]、果实、胚珠和种皮的形成有关[33-37]。
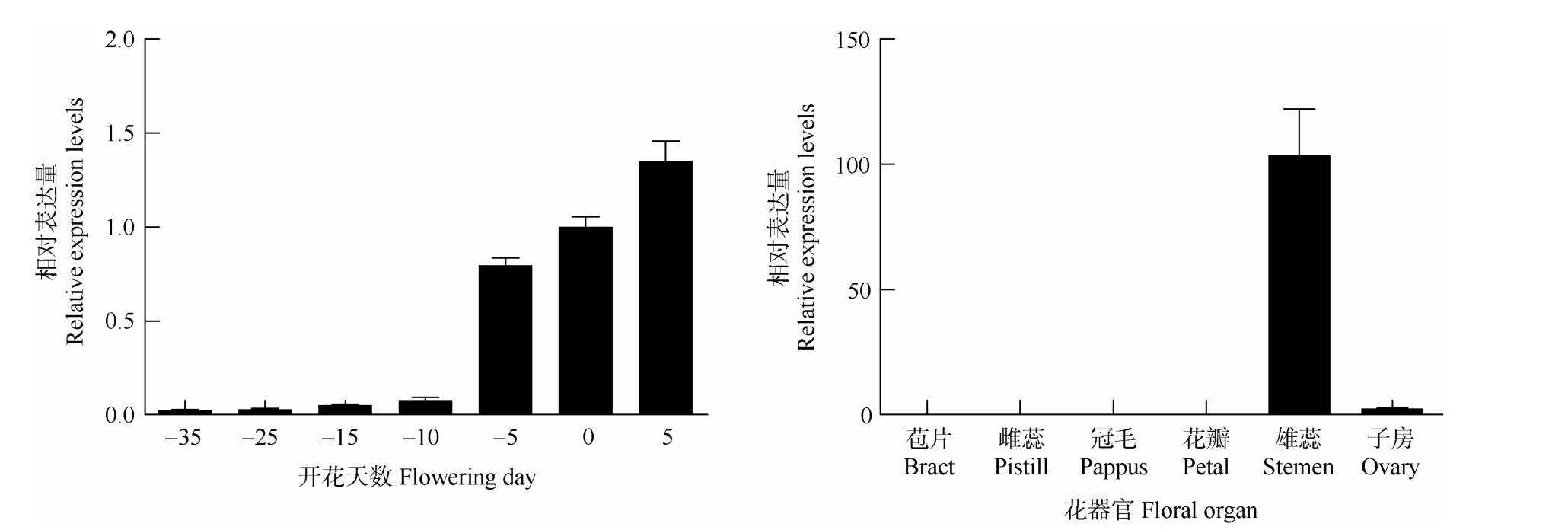
图8 HAM23-like在不同开花时间和不同花器官的表达量Fig.8 Expression of HAM23-like in different flowering day and floral organs
本研究从向日葵中克隆了1个MADS-box基因HAM 23-like,对其编码的蛋白质进行了生物信息学分析,同源对比结果表明HAM 23-like具有典型的MADS-box保守结构域。系统发育分析表明HAM 23-like与拟南芥的AGL18亲缘关系最近。有研究表明;AGL18参与拟南芥花器官发育和角果发育的调控;过表达AGL18会延长生长周期和花器官的衰老和脱落,抑制开花[23]。因此,我们推测向日葵HAM23-like基因作为AGL18的同源基因也可能参与花发育进程调控。AGL18基因在拟南芥营养生长期几乎所有部位有表达,仅新生叶片和下胚轴中没有表达;在生殖生长期从未成熟的花蕾到花粉和种皮均可表达,且其在根、花和角果中的表达量较高[23]。本研究结果表明,HAM23-like基因在向日葵的根、茎、叶、花和瘦果中均有表达,以花和瘦果的表达较高,这与AGL18基因在拟南芥的表达模式较为相似。这进一步说明HAM23-like基因为AGL18的同源基因,也暗示HAM23-like基因可能具有与AGL18基因相似的功能,参与调控向日葵花的形成和开放,而调控方式和调控部位是否相同还有待于进一步研究。
目前,拟南芥和水稻中分别报道了MADS-box基因107和75个[38-43],而有关向日葵的MADS-box基因报道却很少。HAM75和HAM92对花瓣和种皮的形成有影响[17],HAM59和HAM45共同确定雄蕊和雌蕊的形成[18]。但还未见能同时影响雄蕊和果实形成的MADS-box基因。HAM 23-like在花发育过程各阶段均有表达,以开花前5 d、开花当天和开花后5 d的表达量相对较高,且HAM 23-like仅在雄蕊和子房中表达,雄蕊的表达量远高于子房。在拟南芥中AGL18基因未成熟的花蕾、花粉,角果和种皮均具有较高的表达水平[23]。说明向日葵HAM23-like基因也具有拟南芥AGL18基因相似的生物学功能,主要与花发育后期的花器官和花粉发育以及瘦果和种皮早期发育有紧密联系。然而该基因是否在花开放和果实形成过程中起到决定作用还不清楚,而且HAM23-like基因是否和AGL18基因一样对向日葵花器官发育发挥负调控作用也需要进一步的验证[23,44]。
本研究对HAM23-like基因及其编码蛋白的理化性质、结构和功能的预测和分析,以及对该基因在向日葵组织表达的特性进行定量分析,为探究HAM23-like基因在向日葵花发育过程的调控作用提供科学依据。
