稻蟹共作与常规稻田蜘蛛群落组成及多样性分析*
2019-08-06马晓慧车喜庆王井士桑海旭
马晓慧, 车喜庆, 王井士, 桑海旭
稻蟹共作与常规稻田蜘蛛群落组成及多样性分析*
马晓慧, 车喜庆, 王井士, 桑海旭**
(辽宁省盐碱地利用研究所 盘锦 124010)
稻蟹共作是一种新型的稻田生态种养殖技术, 对推动生态农业发展具有重要意义。目前对稻田蜘蛛的研究仅限于常规稻田, 而关于稻蟹共作系统蜘蛛方面的研究尚少见报道。本试验旨在研究稻蟹共作田与常规稻田中蜘蛛群落的组成及多样性特点, 探索稻蟹共作田蜘蛛的发生特点, 为进一步研究稻蟹共作系统生物多样性特征奠定基础。试验采用吸虫器抽吸法, 系统调查了盘锦稻蟹共作田和常规稻田蜘蛛物种组成, 并分析了蜘蛛群落多样性特点。结果表明: 这两类稻田共调查11科31种蜘蛛, 狼蛛科(Lycosidae)、皿蛛科(Linyphiidae)和园蛛科(Araneidae)数量最多, 占总蛛数70%; 稻蟹共作田蜘蛛个体数量显著高于常规稻田。2类稻田蜘蛛优势种均为草间钻头蛛()和拟水狼蛛()。多样性指数表明, 6月初蜘蛛Shannon-Wiener多样性指数最低, 8—9月蜘蛛群落最高; 而均匀度指数、丰富度指数总体与其呈正相关, 优势度指数与其呈负相关; 2类稻田蜘蛛群落多样性特点基本一致。研究结果表明稻蟹共作田和常规稻田蜘蛛种类和多样性特点没有显著差异, 但蜘蛛个体数量有显著差异, 即稻蟹共作田蜘蛛个体数量显著高于常规稻田。
稻蟹共作; 常规稻田; 蜘蛛; 群落; 多样性指数
稻田种养模式是中国生态农业的主要形式之一, 不同地区根据其气候和生产特点形成独具特色的稻田种养模式。我国南方省区主要以稻田养鸭、养虾、养鱼模式为主, 而我国北方则以稻田养蟹模式为主[1-4]。辽宁盘锦位于辽河流域腹地, 水稻()栽培历史悠久, 近年将河蟹养殖与稻作技术结合, 通过减少农药使用, 利用河蟹在稻田的取食与活动来控制稻田有害生物, 形成了典型的稻蟹种养模式。该模式是基于稻作水田条件, 以种稻为中心, 人工投入河蟹苗的自然与人工相配合的生态农业生产技术。
目前关于稻蟹共作技术的研究主要集中于河蟹放养密度、稻蟹田病害与杂草的防治、水稻生长情况及产量等研究, 尚少有稻蟹共作生态系统中节肢动物的研究。节肢动物是生态系统中生物群落的重要组成部分, 通过研究农田生态系统中节肢动物群落, 可以从植物-害虫-天敌相互作用的角度来探讨害虫生态调控的原理和方法, 为科学地控制害虫种群数量提供理论依据[5]。蜘蛛是稻田内仅次于昆虫的种类多、数量大的节肢动物类群, 全部为捕食性, 也是水稻害虫的重要天敌之一[6]。目前我国虽有对棉(spp.)田[7]、稻田[8]、大豆()田[9]及果园[10]等蜘蛛群落结构特征的研究, 但有关稻蟹共作系统蜘蛛群落组成及多样性特点鲜有报道。研究稻蟹共作田与常规稻田蜘蛛群落组成及多样性特点, 有助于探明稻蟹共作生态系统中蜘蛛对有害生物的防控机理及控害程度, 本文系统调查了盘锦单季稻区稻蟹共作田(简称稻蟹田)及常规稻田(简称常规田)蜘蛛群落的组成, 并分析了2类稻田蜘蛛群落多样性特点, 以期为进一步研究稻田种养殖生态系统生物多样性特征、害虫天敌的控害机理以及明确河蟹-植食性昆虫-中性昆虫-捕食性天敌之间营养关系等研究奠定基础。
1 材料与方法
1.1 试验区概况
试验于2018年在辽宁省盐碱地利用研究所试验基地完成, 基地位于辽宁省水稻主产区盘锦市(40°39′~41°27′N、121°25′~122°31′E), 温带季风性气候, 单季稻区, 水稻主栽品种为‘盐丰47’。稻田主要病虫草害为稻瘟病、纹枯病、稻飞虱、二化螟、稻水象甲、稻小潜叶蝇及40余种杂草, 以稗草()、扁秆藨草()、水绵()、鸭舌草()、野慈菇()为优势杂草[11]。稻蟹田仅在5月10日施用恶草灵与丁草胺进行封闭式除草, 之后全年不施化肥与农药, 河蟹投入时间6月1日, 收蟹时间9月24日, 全部为扣蟹, 密度为3 000只∙hm-2, 不投蟹料。常规田于5月9日每块田施30 kg大地丰掺混肥料(锦州市大地丰肥业有限公司, N∶P∶K=28∶18∶8)作为底肥, 5月24日施用丁草胺与吡嘧磺隆防治稗草, 6月2日施用噻虫嗪防治水稻潜叶蝇与稻水象甲, 6月25日施用三唑磷防治1代二化螟, 6月30日施用井冈霉素防治水稻纹枯病, 7月10日施用吡蚜酮防治稻飞虱, 7月20日施用三唑磷防治2代二化螟, 7月28日和8月8日各施用1次吡唑醚菌酯防治稻瘟病; 本田杂草发生重的年份, 需在插秧后再施用五氟磺草胺防治禾本科及莎草科杂草, 其他条件与稻蟹田相同。每块试验稻田面积均为1 000 m2, 周边埝埂种植大豆或留杂草。
1.2 调查时间与方法
水稻5月27日插秧, 10月17日收割。6月1日开始调查, 至10月16日结束, 每月1日、6日、11日、16日、21日、26日调查, 全年共调查28次, 2次重复; 其中稻蟹田最后5次调查时, 田间已没有河蟹(忽略个别漏捕蟹)。蜘蛛调查采用5点法取样, 每样方约1 m2(查4行, 每行6穴), 调查日当天上午用吸虫器在稻株上、中、下3个部位分别抽吸, 并以捕虫网作为补充。蜘蛛样本用75%乙醇保存, 分别记录并带回室内, 参照文献[12-14]清样及鉴定, 统计蜘蛛种类数与个体数, 并将各月调查结果按上、中、下旬统计。本文定义个体数量():>10%为优势种, 5%≤≤10%为丰富种,>1%为常见种。
1.3 数据统计与分析
群落多样性分析以优势度指数()、多样性指数(¢)、均匀度指数()和物种丰富度指数(Ma)表述, 各指数的计算利用Excel 2010完成。具体公式为:
1)优势度分析采用Simpson优势度指数:
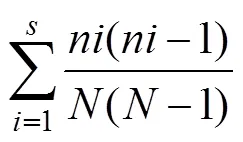
式中:为第个物种的个体数,为所有物种个体数,为物种数。
2)物种多样性分析采用Shannon-Wiener(香农-威纳)多样性指数:
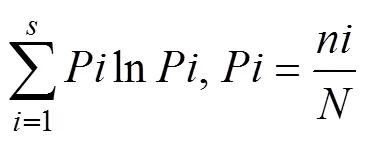
式中:第种个体数占总个体数的比率,为全部物种的个体总数,为物种数。
3)均匀度分析采用Pielou指数:
=¢/max(3)
式中:max=ln,为群落中总的物种数。
4)物种丰富度分析采用Margalef指数:
Ma=(-1)/ln(4)
式中:为物种数,为全部物种的个体总数。
2 结果与分析
2.1 稻蟹田与常规田蜘蛛群落的组成
对稻蟹田和常规田的蜘蛛群落进行调查, 共调查蜘蛛个体样本16 783头, 隶属于11科, 共31种, 详见表1。其中, 稻蟹田共11 747头, 占蜘蛛样本总数量的69.99%, 常规田样本共5 036头, 占蜘蛛样本总数量的30.01%, 可见, 稻蟹田蜘蛛个体数量显著多于常规田。从蜘蛛科来看, 2类稻田蜘蛛个体数量大于15%的均为狼蛛科(Lycosidae)、皿蛛科(Linyphiidae)和园蛛科(Araneidae)。从蜘蛛种来看, 稻蟹田蜘蛛个体总量最多的依次为草间钻头蛛()、拟水狼蛛()、真水狼蛛()、黑斑亮腹蛛()、拟环纹豹蛛()、角类肥蛛(), 常规田蜘蛛个体总量最多的依次为拟水狼蛛、草间钻头蛛、角类肥蛛、黑斑亮腹蛛、真水狼蛛和圆尾肖蛸()。综合来看, 草间钻头蛛和拟水狼蛛为稻田蜘蛛优势种, 真水狼蛛、黑斑亮腹蛛等5种为蜘蛛丰富种, 粽管巢蛛()、黄褐新园蛛()等12种为蜘蛛常见种, 其余为少见或偶见蜘蛛。
蜘蛛物种数随时间变化趋势如图1所示, 稻蟹田和常规田蜘蛛种类随时间变化趋势基本一致, 均由6月初最低值1种, 至8月下旬和9月上旬最高值30种左右, 两种稻田之间无显著差异。
2.2 稻蟹田与常规田蜘蛛群落多样性特征
由于6月上旬采集的稻田蜘蛛种类及数量极少, 2类稻田各参数均为0(图2)。Shannon-Wiener指数从调查开始最小值0, 逐渐增加至9月上旬最大值2.777 5(稻蟹田)和8月下旬最大值2.648 6(常规田), 且8—9月指数均在2.4以上。表明8—9月蜘蛛群落多样性水平高, 物种较丰富, 其他时间多样性水平较低, 尤其是6月初; 稻蟹田和常规田Shannon- Wiener指数变化趋势一致, 无显著差异。
优势度指数从6月初最小值0, 逐渐增加至9月上旬最大值0.912 1(稻蟹田)和8月下旬最大值0.912 3(常规田), 且8—9月指数均保持0.9左右。表明调查初期优势种突出地位并不高, 随着蜘蛛逐渐迁移至稻田内, 优势种突出地位也越来越明显, 直到8月底9月初, 优势种的突出地位最明显, 即草间钻头蛛和拟水狼蛛在此时优势地位突出。稻蟹田和常规田优势度指数变化趋势一致, 无显著差异。
均匀度指数在6—7月稍有波动, 从最小值0到6月中旬最大值0.999 9(稻蟹田)和7月上旬最大值0.881 6(常规田), 其余阶段波动不大, 指数值均以0.7~0.8为主。均匀度指数间接地反映蜘蛛群落中优势种的突出程度, 可见, 优势种蜘蛛突出程度在8—9月变化不大, 即两者以一定的种群数量始终保持着一定的突出地位。
物种丰富度指数变化趋势与Shannon-Wiener指数基本一致, 但稻蟹田和常规田稍有差异, 指数最低值在6月上旬, 均为0, 指数最高值不同, 稻蟹田出现在7月下旬, 为4.426 7, 常规田出现在8月中旬, 为4.181 7。且6—8月稻蟹田物种丰富度指数略高于常规田, 9月起反之。这表明稻蟹田7月下旬蜘蛛物种数量最多, 之后物种数逐渐减少, 而常规田8月中旬后, 蜘蛛物种数才开始减少。
Shannon-Wiener指数随时间的变动规律, 反映出蜘蛛种群从迁入稻田至水稻近收割期间群落多样性水平整体的变动趋势。当蜘蛛群落多样性程度较高的阶段, 均匀度和物种丰富度程度相应的也较高, 而优势度指数则较低, 即优势种在群落中突出地位越不明显。这也说明, 各参数间具有一定的相关性, 均匀度指数、丰富度指数与Shannon-Wiener指数呈正相关性, 而优势度指数与其呈负相关性。稻蟹田与常规田多样性指数变化趋势基本一致, 仅在物种丰富度指数上稍有差异, 这可能受河蟹捕食与活动影响, 具体原因有待进一步研究。
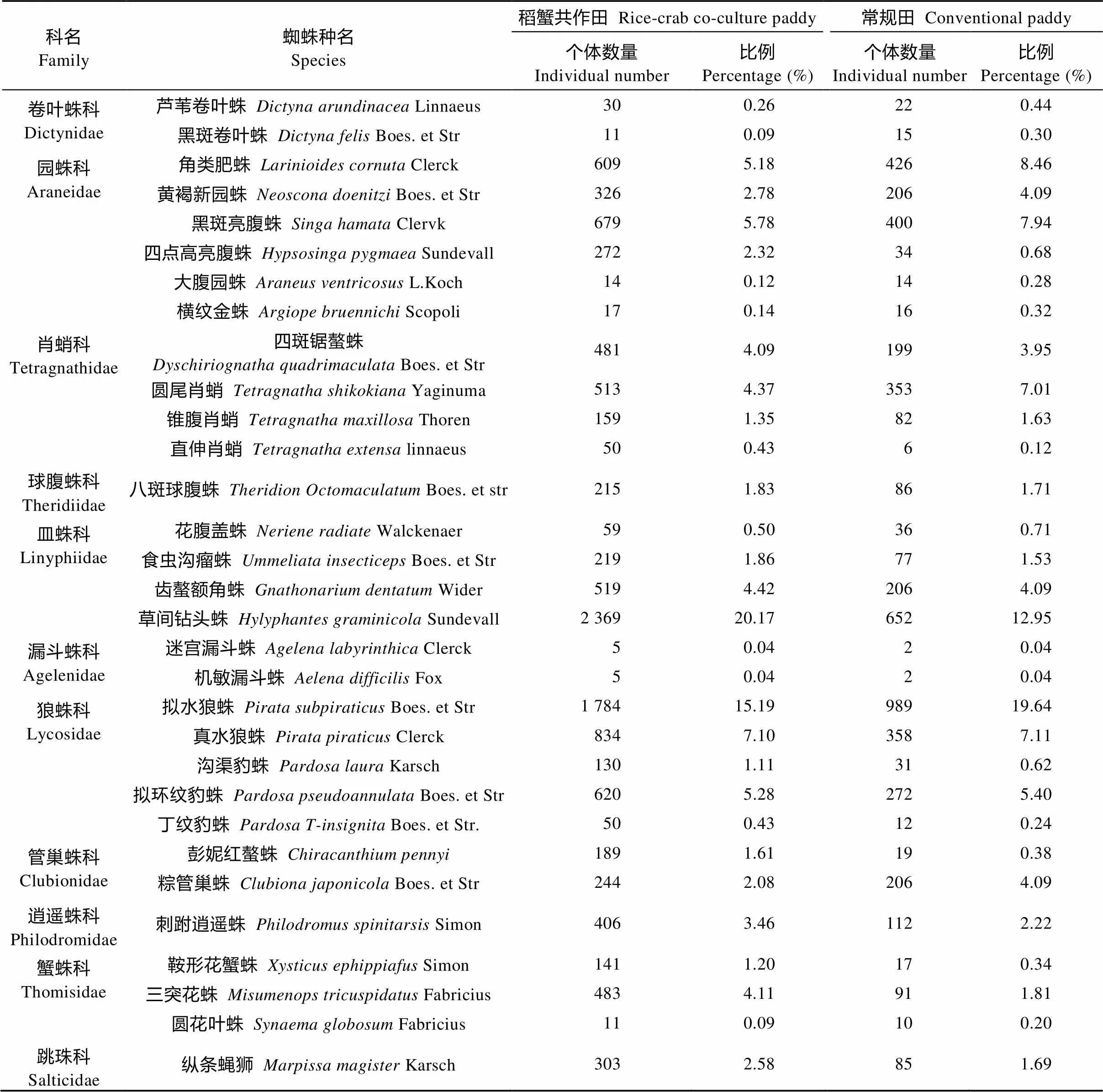
表1 稻蟹共作田和常规稻田蜘蛛群落组成
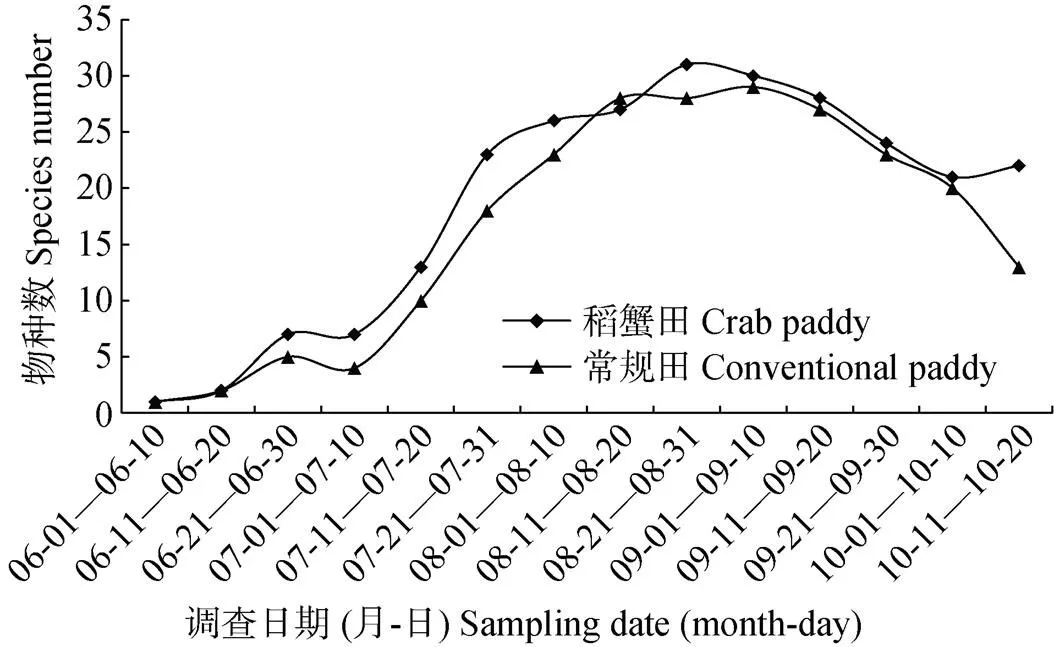
图1 稻蟹共作田和常规稻田蜘蛛物种数随时间的变化
3 讨论与结论
稻蟹共作作为一种新型养殖模式, 在经济效益、社会效益和生态效益方面较常规水稻栽培模式具有较大优势[15]。稻蟹共作模式主要集中在我国北方沿海地区, 据2006年统计, 辽宁省河蟹蟹种生产量达19 020 t, 居全国首位[4]。针对稻蟹共作生态系统, 前人主要开展了杂草生态防控[16-17]、稻瘟病药剂防治[18]、药物安全使用[19]以及河蟹放养密度及水稻栽培模式对水稻和河蟹的影响等相关研究[15,20], 尚少有对稻蟹共作生态系统蜘蛛种群多样性特点的研究。本文调查结果表明, 稻蟹田和常规田蜘蛛种类丰富, 共调查蜘蛛标本16 783头, 隶属于11科, 共31种, 以狼蛛科、皿蛛科、园蛛科蜘蛛数量最多, 稻蟹田蜘蛛个体数量显著多于常规田。2类稻田蜘蛛优势种均为草间钻头蛛和拟水狼蛛, 这与王井士等[21]对盘锦稻区蜘蛛优势种调查结果略有不同, 这是因为本文调查仅限在水稻植株上采集, 不包含周边非稻田生境。2类稻田中蜘蛛优势种略有不同, 如稻蟹田常见种圆尾肖蛸个体数占4.37%, 而常规田该种类为丰富种, 占7.01%, 圆尾肖蛸在稻株之间结大型水平圆网, 居于水稻中上部, 蜘蛛在网附近稻株上活动, 而稻株中部叶鞘等处也是河蟹活动的范围, 这是否与河蟹取食、活动有关, 还有待进一步研究。蜘蛛多样性指数分析表明, 6月初蜘蛛群落多样性水平最低, 8—9月最高, 而均匀度、丰富度指数总体与其呈正相关, 优势度指数与其呈负相关, 稻蟹田与常规田多样性指数变化趋势基本一致, 这与徐雪亮等[22]2018年对施药稻田和不施药稻田蜘蛛群落多样性、均匀度、优势度均无显著差异的研究结果一致, 稻蟹田为不施药稻田, 推测蜘蛛群落多样性受稻田施药的影响不大。
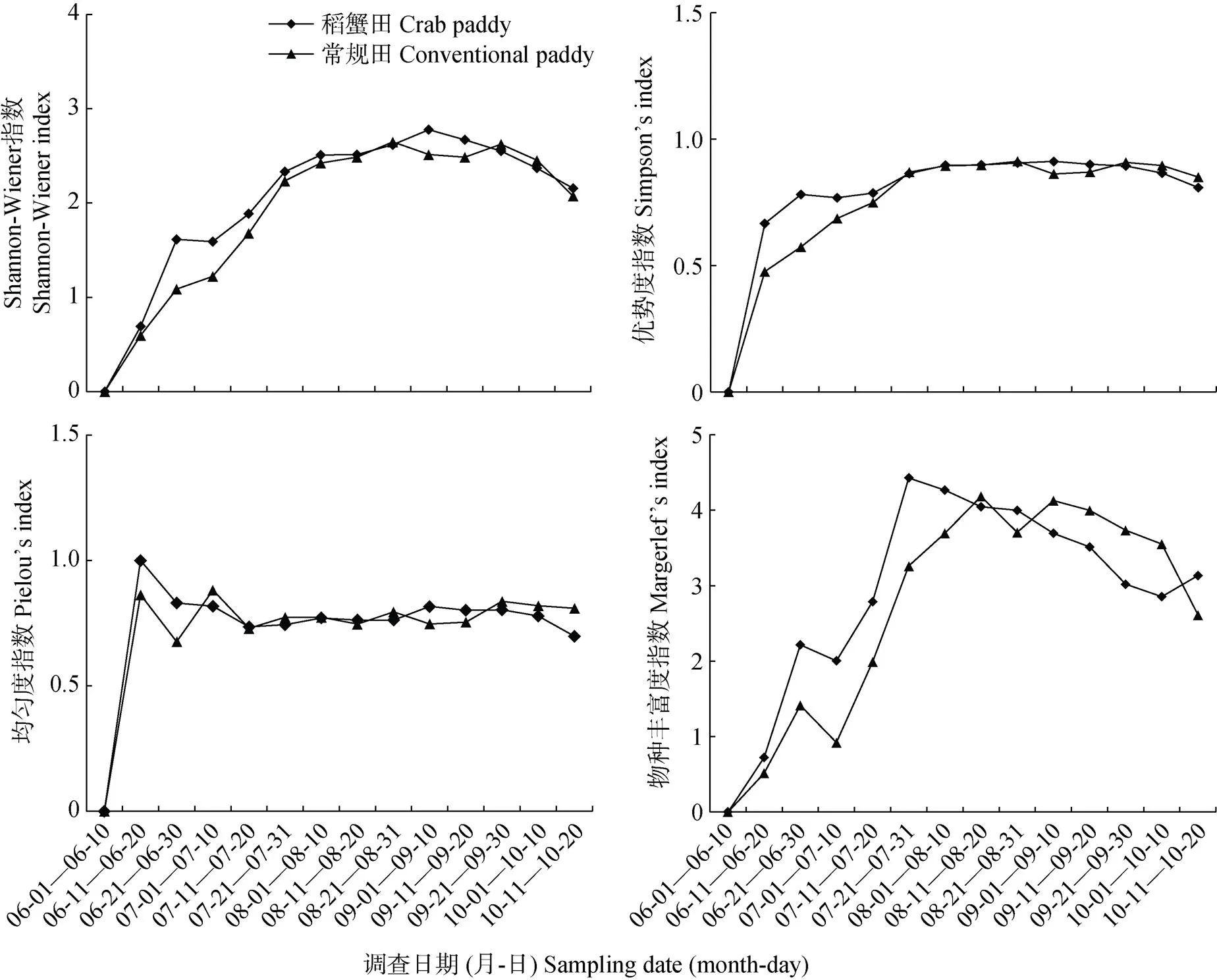
图2 稻蟹共作田和常规稻田蜘蛛多样性随时间的变化规律
Table 2 Time distribution of spider diversity in rice-crab co-culture paddy and conventional paddy
本文研究表明, 常规田因为不间断使用化学药剂, 使得害虫与天敌数量大量减少, 群落重建速度慢, 生物量减少使得生物多样性程度也降低, 因而天敌的控制能力有限。稻蟹田由于不施用化学药剂, 即便河蟹取食一定量害虫与天敌, 稻蟹田生物量也显著高于常规田。另外, 河蟹在稻田捕虫、啃草、掘土等取食与活动, 间接起到除草、防虫、净土的作用, 对水稻生长有一定的促进作用。本文对稻蟹共作田蜘蛛群落组成进行研究, 明确了蜘蛛种类与数量, 为后续研究奠定基础。但当稻田植入河蟹这一生物, 并取消施用农药之后, 稻田生态系统生物多样性发生了变化, 河蟹与稻田中植食性、捕食性和中性节肢动物间相互作用关系仍有待明确, 有关各类蜘蛛捕食力、蜘蛛对稻田有害生物控制力、蜘蛛生态位情况等, 尚需进一步研究。
[1] 张林林. 稻田养鱼技术模式演变及发展趋势分析[J]. 现代农业科技, 2007, (18): 160–161 ZHANG L L. Rice-fish technology schema evolution and development trend analysis[J]. Modern Agricultural Science and Technology, 2007, (18): 160–161
[2] 任志如. 龙虾稻田养殖技术[J]. 水产养殖, 2012, 33(3): 22–25 REN Z R. Lobster paddy field aquaculture technology[J]. Journal of Aquaculture, 2012, 33(3): 22–25
[3] 禹盛苗, 金千瑜, 朱练峰, 等. 稻田养鸭密度对水稻产量和经济效益的影响[J]. 浙江农业科学, 2008, (1): 68–71 YU S M, JIN Q Y, ZHU L F, et al. Effects of ducks density in rice paddy on rice yield and economic benefit[J]. Journal of Zhejiang Agricultural Sciences, 2008, (1): 68–71
[4] 王武. 北方稻田养蟹产业发展思路[J]. 中国水产, 2008, (10): 11–13 WANG W. North rice paddy industry development way[J]. China Fisheries, 2008, (10): 11–13
[5] 史树森, 高月波, 臧连生, 等. 不同杀虫剂对大豆田节肢动物群落结构的影响[J]. 应用昆虫学报, 2012, 49(5): 1249–1254 SHI S S, GAO Y B, ZANG L S, et al. Effects of several insecticides on the arthropod community in soybean fields[J]. Chinese Journal of Applied Entomology, 2012, 49(5): 1249–1254
[6] 马晓慧, 桑海旭, 刘郁. 辽宁盘锦稻区杂草地与稻田中蜘蛛动态[J]. 植物保护, 2015, 41(3): 165–168 MA X H, SANG H X, LIU Y. Fluctuation of spiders in paddy field and adjacent weed habitat in Panjin, Liaoning Province, China[J]. Plant Protection, 2015, 41(3): 165–168
[7] 上官小霞, 沈文君, 李生才. 棉田蜘蛛群落时空生态位研究[J]. 中国生态农业学报, 2002, 10(4): 87–90 SHANGGUAN X X, SHEN W J, LI S C. Study on the temporal-spatial niche of spider community in cotton field[J]. Chinese Journal of Eco-Agriculture, 2002, 10(4): 87–90
[8] 马晓慧, 桑海旭, 王井士, 等. 盘锦稻区蜘蛛群落的建立与多样性分析[J]. 植物保护, 2017, 43(6): 168–172 MA X H, SANG H X, WANG J S, et al. Community establishment of spiders and diversity analysis in paddy field of Panjin[J]. Plant Protection, 2017, 43(6): 168–172
[9] 高月波, 史树森, 孙嵬, 等. 大豆田节肢动物群落优势种群时间生态位及营养关系分析[J]. 应用昆虫学报, 2014, 51(2): 392–399 GAO Y B, SHI S S, SUN W, et al. Analysis of the temporal niches of dominant species and nutritional relationships within the arthropod community in soybean fields[J]. Chinese Journal of Applied Entomology, 2014, 51(2): 392–399
[10] 李生才, 高峰, 王宁波, 等. 苹果园蜘蛛群落组成及其生态位研究初报[J]. 中国生态农业学报, 2006, 14(1): 181–184 LI S C, GAO F, WANG N B, et al. Composition and niche of spider community in apple orchard[J]. Chinese Journal of Eco-Agriculture, 2006, 14(1): 181–184
[11] 马晓慧, 王井士, 桑海旭, 等. 辽宁水稻主产区杂草种类调查及其优势度[J]. 北方水稻, 2018, 48(5): 22–25 MA X H, WANG J S, SANG H X, et al. Weeds species investigation and the dominance in main rice producing area of Liaoning Province[J]. North Rice, 2018, 48(5): 22–25
[12] 陈孝恩, 高君川. 四川农田蜘蛛彩色图册[M]. 成都: 四川科学技术出版社, 1990: 41–195 CHEN X E, GAO J C. Color Atlas of Farmland Spider in Sichuan[M]. Chengdu:Sichuan Science and Technology Press, 1990: 41–195
[13] 何俊华, 庞雄飞. 水稻害虫天敌图说[M]. 上海: 上海科学技术出版社, 1986: 208–235 HE J H, PANG X F. Natural Enemies of Rice Pests[M]. Shanghai:Shanghai Science and Technology Press, 1986: 208–235
[14] 张志升, 王露雨. 中国蜘蛛生态大图鉴[M]. 重庆: 重庆大学出版社, 2017 ZHANG Z S, WANG L Y. Chinese Spiders Illustrated[M]. Chongqing: Chongqing University Press, 2017
[15] 徐敏, 马旭洲, 王武. 稻蟹共生系统水稻栽培模式对水稻和河蟹的影响[J]. 中国农业科学, 2014, 47(9): 1828–1835 XU M, MA X Z, WANG W. Effects of different cultivation patterns on rice yield and crab in rice-crab culture system[J]. Scientia Agricultura Sinica, 2014, 47(9): 1828–1835
[16] 吕东锋, 王武, 马旭洲, 等. 稻蟹共生对稻田杂草的生态防控试验研究[J]. 湖北农业科学, 2011, 50(8): 1574–1578 LYU D F, WANG W, MA X Z, et al. Ecological prevention and control of weeds in rice-crab polycultured field[J]. Hubei Agricultural Sciences, 2011, 50(8): 1574–1578
[17] 于凤泉, 孙富余, 李志强, 等. 稻蟹生态种养田水绵防除药剂的筛选[J]. 辽宁农业科学, 2011, (4): 75–76 YU F Q, SUN F Y, LI Z Q, et al. Drug screening for spirogyra control in rice-crab culture system[J]. Liaoning Agricultural Sciences, 2011, (4): 75–76
[18] 于凤泉, 李志强, 赵旭, 等. 不同药剂对稻蟹生态种养田水稻稻瘟病的防治效果研究[J]. 辽宁农业科学, 2013, (1): 78–79 YU F Q, LI Z Q, ZHAO X, et al. Study on the control effect of different chemicals on rice blast in rice-crab paddy[J]. Liaoning Agricultural Sciences, 2013, (1): 78–79
[19] 龚蕾蕾, 范丽萍, 高峰, 等. 稻蟹种养模式中水稻药物的安全使用技术[J]. 水产科技情报, 2010, 37(4): 202–203 GONG L L, FAN L P, GAO F, et al. Secure application of drugs in mixed culture of crab and rice[J]. Fisheries Science & Technology Information, 2010, 37(4): 202–203
[20] 闫志利, 林瑞敏, 牛俊义, 等. 我国稻蟹共作技术研究的现状与前景展望[J]. 北方水稻, 2008, 38(2): 5–8 YAN Z L, LIN R M, NIU J Y, et al. Current status and prospectives of rice-crab production technique research in China[J]. North Rice, 2008, 38(2): 5–8
[21] 王井士, 马晓慧, 桑海旭. 辽河三角洲稻区优势天敌种类调查分析[J]. 植物保护, 2015, 41(1): 163–165 WANG J S, MA X H, SANG H X. Investigation and analysis of dominant natural enemies in Liaohe River Delta[J]. Plant Protection, 2015, 41(1): 163–165
[22] 徐雪亮, 刘子荣, 王奋山, 等. 不同防治措施对江西稻田蜘蛛亚群落结构的影响[J]. 中国农学通报, 2018, 34(1): 118–123XU X L, LIU Z R, WANG F S, et al. Effects of different control measures on spider subcommunity structure in rice field[J]. Chinese Agricultural Science Bulletin, 2018, 34(1): 118–123
The structure of spider communities in crab paddies and conventional paddies*
MA Xiaohui, CHE Xiqing, WANG Jingshi, SANG Haixu**
(Liaoning Saline or Alkaline Land Utilization and Research Institute, Panjin 124010, China)
Rice-crab co-culture (RC) is a new eco-agricultural process in paddy cultivation and breeding models. It has been significantly promoted in the development of eco-agricultural programs. Recent studies, which concern the RC system, mainly focus on the effects of multiple factors on crab or rice production. However, studies on the community of natural enemies in RC ecosystems are still rare. The goal of this study was to determine the regulation of occurrence, composition, and diversity of spider communities in crab paddies and conventional paddies. Suction samplers were applied to collect spiders in the paddies. Species composition and diversity of spiders were recorded and analyzed.The results showed that: 1) 31 species and 11 families of spiders were found in both paddies. Three major families of spiders,,, and, accounted for at least 70% of the spider populations. 2) The number of individual spiders in the crab paddies was significantly higher than that in conventional paddies. 3) The Shannon-Wiener index indicated that the diversity of spider species was highest from August through September, and lowest in early June. Furthermore, the results showed that the Shannon-Wiener indexes were positively correlated with Pielou’s or Margalef’s indexes but were negatively correlated with the Simpon’s indexes. The results revealed that there was no difference in the numbers or diversities of spider species between crab paddies and conventional paddies, whereas the difference in the number of individual spiders was significant between the two paddy types. This study lays a foundation for the study of the biodiversity characteristics of arthropods in RC systems.
Crab paddy; Conventional paddy; Spider; Community; Diversity index
S476
2096-6237(2019)08-1157-06

10.13930/j.cnki.cjea.181068
* 国家重点研发计划项目(2018YFD0200203, 2016YFD0300703)资助
桑海旭, 主要研究方向为水稻病虫害防治。E-mail: haixusang@163.com
马晓慧, 主要从事水稻病虫草害综合防治研究。E-mail: jodiexiaohui@163.com
2018-12-08
2019-04-08
* The work was supported by the National Key Research and Development Project of China (2018YFD0200203, 2016YFD0300703).
, E-mail: haixusang@163.com
Dec. 8, 2018;
Apr. 8, 2019
马晓慧, 车喜庆, 王井士, 桑海旭. 稻蟹共作与常规稻田蜘蛛群落组成及多样性分析[J]. 中国生态农业学报(中英文), 2019, 27(8): 1157-1162
MA X H, CHE X Q, WANG J S, SANG H X. The structure of spider communities in crab paddies and conventional paddies[J]. Chinese Journal of Eco-Agriculture, 2019, 27(8): 1157-1162
