平面细胞极性通路在神经管缺陷中的作用
2019-08-05黄少勇蔡润泽陈永昌
黄少勇,蔡润泽,陈永昌*
(1.昆明理工大学灵长类转化医学研究院,昆明 650500; 2.云南中科灵长类生物医学重点实验室,昆明 650500)
神经管缺陷也被称作神经管畸形(neural tube defects,NTDs),其发病率仅次于先天性心脏病,是最常见先天性畸形[1]之一,通常是由于胚胎发育早期神经管闭合不完全或者不闭合造成的。NTDs一旦发生则不可逆转,多在孕期流产或出生后很短的时间内死亡[2]。目前,尽管已有神经管缺陷的产前诊断方法,但是全球每年有超过30万的NTDs婴儿出生,而患有NTDs的婴儿有较高的死亡率,十分之一患有NTDs的婴儿会在第一年死亡[3]。神经管畸形可分为无脑儿(anencephaly)、脑膨出(encephalocele)、脊柱裂(spina bifida)和颅脊柱裂(craniorachischisis)等几种类型。NTDs在全世界范围内的发病率已经达到0.5‰~2‰[4],并且在不同的地区发病率也有着较大的差别。神经管闭合依赖于肌球蛋白收缩的细胞运动过程例如根尖收缩,组织和细胞定向重排等过程[5]。NTDs的发生不仅受遗传因素影响,也受到环境因素共同影响,目前常用补充叶酸(folic acid,FA)的方法来预防神经管缺陷的发生,妊娠前和妊娠早期补充适量的叶酸可以降低60%~70%的NTDs的发生[6]。此外,体外胚胎干细胞分化过程中也发现叶酸对神经分化具有促进作用[7]。因此,在对神经管畸形的发生及其致病机制的研究中,大部分研究的焦点主要集中在叶酸代谢通路及其相关基因,但补充叶酸只能预防NTDs并不能阻止它的发生且叶酸预防NTDs发生的具体分子机制尚不清楚[7]。了解研究神经管缺陷的致病机制有助于人们对NTDs的治疗。近年来科学家通过对小鼠、非洲爪蟾等神经管畸形动物模型的研究表明,平面细胞极性(planar cell polarity,PCP)信号通路在神经管闭合的过程中起着很重要的作用,PCP信号通路及其相关基因也成为研究神经管畸形遗传基础的热点之一。因此,建立合适的动物模型对了解神经管缺陷的致病机制,进而更好地研究预防和治疗NTDs至关重要。
1 PCP信号通路
神经胚的形成是胚胎发育中的一个基本过程。神经胚的发育影响神经管的发育,而大脑和脊柱的前体是神经管[8]。神经管是在胚胎发育过程中由大脑和脊髓的中枢神经形成的一个瞬态结构[7]。神经管的闭合是由神经板的折叠、升高、结合、在背侧中线处融合,从而形成神经管[9]。在这一过程中的任何一个步骤发生中断就会导致NTDs,例如脊柱裂、无脑儿、无脑畸形和颅脊柱裂[7]。因此在神经管闭合的过程中每一步都显得至关重要,而在整个过程中PCP信号通路有着关键作用。PCP信号通路也被称为非典型Wnt信号通路,它调节与基底垂直的上皮片的极性[10]。最早在果蝇中发现PCP信号通路,其影响果蝇翅膀体毛排列和复眼等高度有序化结构的形成[11]。在哺乳动物中,节点纤毛的定位以及内耳的纤毛和皮肤上的毛囊中都发现了PCP信号通路的存在[12]。Adler等人[13]的研究表明,PCP信号通路在进化上非常保守。PCP核心蛋白会形成一个比较复杂的膜复合体,这些蛋白质的数量和亚细胞分布对胚胎发育中原肠胚的形成以及神经胚形成过程中的PCP信号通路成员的汇聚延伸(convergent extension,CE)发挥着重要的作用[14]。CE是指在原肠胚形成的后期,神经管在后脑与颈部交界的区域闭合,神经管的闭合要求底板细胞群向中线汇聚延伸的一个过程[15]。CE是由非典型Wnt/Frizzled信号通路控制。与典型Wnt信号通路不同的是,非典型Wnt信号通路是通过β-actin稳定的控制细胞的命运,即果蝇中PCP通路[16],而且CE这一过程的发生是由细胞的极化重排驱动的[17]。因此PCP通路驱动CE让细胞发生重新形成神经板,SHROOM3、VANGL2和WNT5A基因通过改变细胞形态使神经板折叠直至神经管闭合形成。
2 PCP通路的相关基因
PCP通路包括以下6个核心基因:FRIZZLED(FZ),DISHEVELLED(DSH/DVL),STRABISMUS/VANGOGH(STAM/VANGL),FLAMINGO(FMI)/CELSR,PRICKLE(PK)和DIEGO(DGO)等[20]。果蝇FZ基因在多年前就被发现能调节平面极性[21],而且FZ蛋白参与了几乎所有的Wnt信号通路。但不同的FZ蛋白参与不同的通路,在中枢神经中神经轴突的生长以及方向定位中就有FZD3蛋白在发挥着作用[22]。DSH基因是Wnt信号通路的关键组成部分,可能也有其他的功能,DSH是Wnt信号的关键因素,这些信号来源于Wnt受体家族,即卷曲蛋白。DSH能指挥来自于细胞质膜的信号进入正确的细胞内通路[23]。VANGL基因是PCP通路中第一个确定与人类NTDs有关的基因,并且有VANGL1和VANGL2两个亚型。非洲爪蟾同源人类VANGL1和VANGL2负性调节Wnt β-catenin信号通路,编码Wnt受体β信号通路负调控基因的功能突变会导致肿瘤发生[24]。FMI基因也是在果蝇中发现的,并且与PCP通路相关,FMI基因在果蝇的神经系统发育中起着很重要的作用[25]。Chen 等人[26]对184例中国NTDs病人的基因分析,他们发现3个CELSR基因(CELSR1,CELSR2 和CELSR3)发挥重要作用,这3个基因的突变会导致NTDs患病的风险大大增加。PK基因在细胞层影响VANGL1的不对称动力学,因此,PK基因控制自己的定位与VANGL1产生共同的影响[27]。DGO蛋白在脊椎动物上的同源物是DVL,而DVL与Wnt信号通路和非经典Wnt信号通路都有关[28],目前在NTDs上研究不多,这些基因都是与PCP信号通路有关。还有一些基因在神经管闭合这一过程中也起着比较重要的作用,例如SHROOM3和肌醇代谢中的PIP5K1等基因,Wang等[7]发现在PIP5KL缺失的小鼠中,神经管闭合有缺陷,而且神经上皮排列混乱。
3 PCP信号通路与NTDs
已知PCP信号通路核心基因对NTDs的重要性,但除此之外,还有许多基因也对神经管的闭合有着很重要的作用,例如:SHROOM3和肌醇代谢中的PIP5K1等基因,Wang等人[7]发现在PIP5KL缺失的小鼠中,神经管闭合有缺陷,而且神经上皮排列混乱。下面对SHROOM3,VANGL2和WNT5A这三个基因是如何影响PCP信号通路,进而导致NTDs的发生进行阐述。
Jeffrey等[29]对小鼠的SHROOM3基因研究发现,SHROOM3基因的严重突变会导致严重的大脑畸形。因此,SHROOM3基因可能对神经管闭合过程中是一个关键的因素。在大多数脊椎动物中它SHROOM3基因的正常表达是神经管形成的必要因素,该基因在进化上是保守的。在该研究中,SHROOM3基因的功能的丧失会导致小鼠发生无脑畸形、脑膨出、脊柱裂等畸形,说明神经管闭合失败。其原因是SHROOM3的功能丧失会使神经折叠结构向外部扩展而不能在背侧中线处汇合成。SHROOM3编码一个PDZ结构域的蛋白质,并且从多层次来调节细胞结构。首先,内源性SHROOM3蛋白黏附于细胞骨架上;第二,局部表达的SHROOM3蛋白改变纤维肌动蛋白的分布,并且该蛋白质可以与纤维肌动蛋白结合;第三,SHROOM3蛋白直接结合肌动蛋白;最后,神经上皮的细胞骨架会影响突变体的胚胎[30]。SHROOM3基因的缺失会阻断神经胚的形成,导致神经管闭合失败,由此引发的神经管闭合缺陷并不能通过叶酸补充来预防,反而大剂量的补充会增高死亡率。目前,SHROOM3基因在胚胎发育过程中发挥功能的机制尚不清楚。除了SHROOM3基因之外,Chen和他的团队[31]对NTDs的病人研究发现,SHROOM2基因的突变会破坏PCP信号通路,进而改变细胞骨架后重塑导致NTDs风险增加。
VANGL2基因是1988年Taylo[32]在从果蝇的基因组发现的,根据表型不同,最先发现的基因而被命名为STBM或VANGL2。随着研究的深入,Kibar和他的同事[33]推测人类的NTDs由VANGL1及VANGL2基因突变引起的,这两个基因在小鼠中与平面细胞极性有关。在NTDs患者中发现了很多种VANGL2基因突变,证实了NTDs的发生与VANGL2基因有密切的关系[34]。在脊椎动物的早期胚胎发育过程中,CE驱动胚胎轴和神经板的收缩和延长。在几种模式生物的原肠胚中,VANGL2在CE过程中起着决定性的作用[35]。Park等人[36]发现在爪蟾胚胎中过表达VANGL2能够抑制CE的发生。如果在这一过程中VANGL2基因发生严重突变,可能导致高度组织特异性的上皮结构失去正常的极性,从而导致不同程度的神经管缺陷的发生。Gabriel L Galea和他的团队[37]在VANGL2突变的纯合小鼠中发现,VANGL2的突变阻止了小鼠神经管的正常闭合,导致了NTDs的发生。因此,阐明VANGL2基因在神经管闭合的过程中的分子作用机制,才能有效的预防或者阻止NTDs的发生。
1999年,Yamaguchi等[38]发现WNT5A基因的缺失会导致胚胎在发育过程出现差错从而导致新生儿的死亡。Shimizu[39]发现,在果蝇中WNT基因作用于PCP信号通路的上游。Wnt信号通路非常保守,且与神经管畸形密切相关。Kimura-Yoshida等[7]发现Wnt信号调制器DKK1和KREMEN1定位在神经板边界。Kilian等人[40]发现WNT5A是后内胚层如脊索中CE过程所需要的。Mcgreevy等[41]发现WNT5A是PCP通路中的一个配体,在WNT5A敲除的小鼠中呈现特有的CE过程,WNT5A和SHROOM3在遗传上是相互作用的。2017年对小鼠的研究发现,PCP通路的一些组分会调节WNT5A,使皮质脊髓轴突成长成圆锥细胞[5]。
4 NTDs动物模型
NTDs的致病原因相当复杂,与单基因控制的疾病不同,遗传因素和环境因素共同作用于NTDs。在遗传因素中,虽然发现了许多基因与神经管闭合有关,但是这些基因具体如何发挥作用导致NTDs尚不清楚。建立合适的动物模型是探究相关基因及信号通路、营养因素等与NTDs相关性的重要手段。
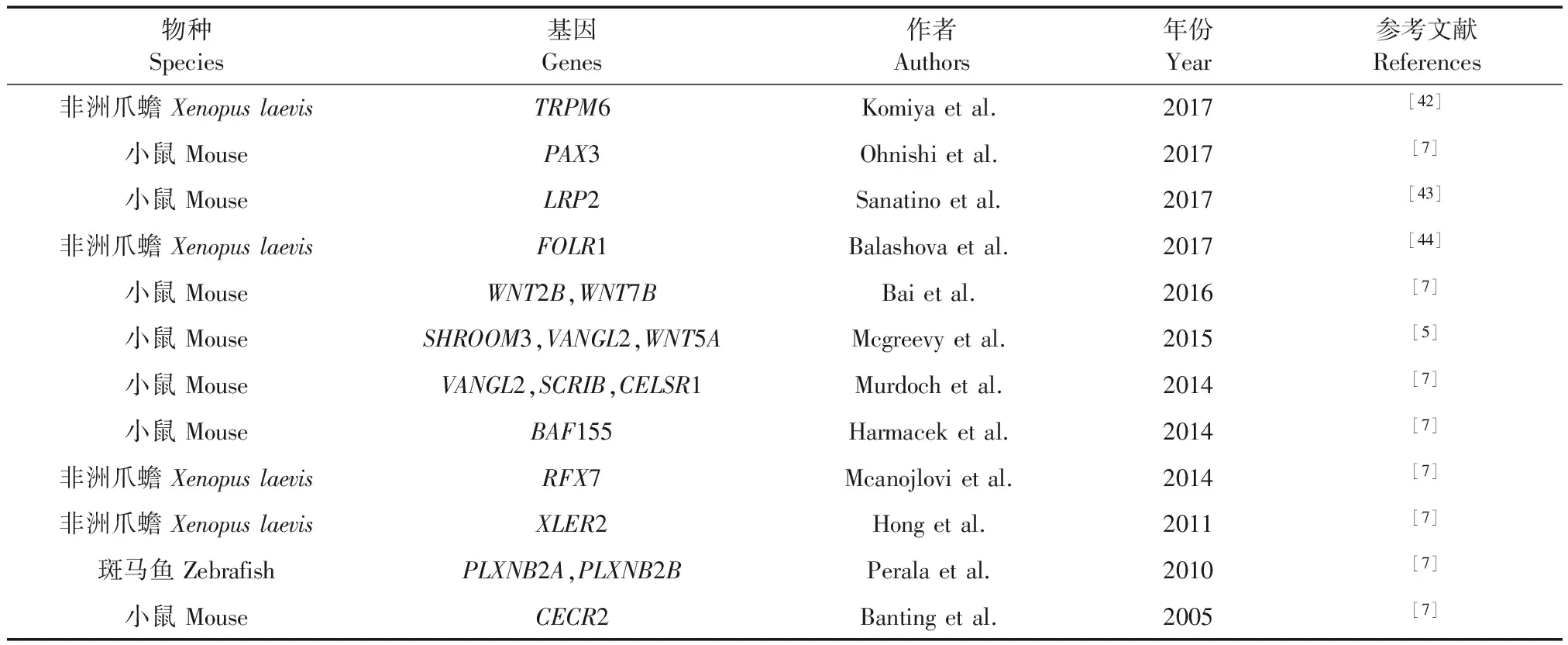
表1 主要NTDs动物模型及相关基因
有关NTDs代表性动物研究模型构建见表1。从表中我们可以得出目前大部分研究NTDs的动物模型都集中在小鼠、非洲爪蟾和斑马鱼等低等模式动物。因为NTDs的发生是在神经发育早期发生的,用其他的模式动物很难观察到神经管闭合的过程。而上述三种动物模型则可以很清楚和直观的观察到神经管闭合过程中的变化,并且敲除这些动物的与NTDs相关的基因很容易实现,因此很多科学家用这些动物来研究NTDs。在这些研究模型中,很多研究的基因是PCP信号通路中的主要基因,也有其它一些重要基因如SHROOM3,TRPM6,和PAX3等基因。在表中的这些基因都与神经管闭合的过程中的PCP信号通路以及CE过程有着很重要的关系。由于基因编辑技术的发展,这些动物模型的建立更加容易,但是也存在一些局限性。小鼠、非洲爪蟾和斑马鱼的大脑发育有着很大的差别,一些基因表达的时间和空间也不尽相同。对于人类NTDs来说,这些模式动物的大脑发育和人类大脑发育有着天壤之别。在这些模式动物中,某些与NTDs相关基因的突变可以导致它们发生神经管闭合不全,但是这些结果不一定在人类上可以发生。由于非人灵长类动物的大脑和人发育的相似度很高,所以研究人类疾病的动物模型非人灵长类动物是最好的选择。但是由于嵌合体的问题,在非人灵长类动物上产生人类NTDs有着很大的难题。因此,研究与人类大脑发育及其相似的非人灵长类动物模型迫在眉睫。
5 问题与展望
本文通过SHROOM3,VANGL2和WNT5A三个基因来阐明PCP信号通路在神经管缺陷中的一些作用和和分子机制。在正常的发育过程中,细胞表达SHROOM3蛋白,产生的蛋白使细胞呈楔型,并且变长,引起神经板卷曲。还有一些细胞如附近的皮肤和表皮细胞等,则推动和形成折叠,从而促使神经管的闭合。VANGL2基因参与了CE这个过程,而CE又是PCP信号通路的一个重要组成部分,因此VANGL2基因的突变也会导致神经管的闭合不全。而WNT5A基因作用与PCP信号通路的上游来参与神经管闭合这一过程,但是WNT5A基因突变不一定会导致神经管缺陷的发生。
目前世界各国采用给育龄妇女补充FA来预防NTDs的发生,虽然达到了一定的预防效果,但还是不能阻止NTDs的发生。问题在于NTDs的致病原因复杂,而且与之相关的基因较多。补充叶酸可以预防由VANGL2基因突变导致的NTDs,但是还有很多基因的突变导致的NTDs是叶酸所不能预防的。近年来,很多科学家都投入到研究NTDs的工作中,虽然取得了很大的成功,但是离治疗还有很大一段路要去探索。目前研究的NTDs的动物模型只有小鼠,大鼠和斑马鱼等模式生物,这些动物模型只能帮助了解和探索NTDs的发病机制和作用机理。但是,这些动物模型与人类的发育有着很大的差别,因此建立非人灵长类NTDs模型至关重要,而且一个成功的NTDs非人灵长类动物模型对发现新的治疗方法和预防NTDs的发生有着十分重要的意义。
