独行菜族8属(十字花科)植物果实及种子微形态研究
2019-07-23曹董玲张学杰
曹董玲 张学杰* 刘 玫
(1.山东师范大学,济南 250014; 2.哈尔滨师范大学,哈尔滨 150025)
独行菜族(Trib. Lepidieae)为十字花科(Cruciferae)较大类群,因其族内属间具有稳定丰富的性状,长期以来被公认为自然类群。我国有20属,约48种、10变种和1变型[1~2]。植株常为草本,稀为半灌木,果实为角果,开裂,少数不裂。该族部分植株全草具有较高的科研和药用价值,相关研究方向涉及广泛[3~7]。
近年来随着分子系统学、化学成分分析及耐逆胁迫机制相关研究报道逐渐深入,在利用核基因和序列进行亲缘关系演化分析中,结果与传统分类系统研究存在一定分歧且缺乏相关微形态结构依据[8~12]。独行菜族内部分属、种的分类鉴定仍存在一定困难及争议,部分属、种界限尚不明显,对于十字花科果实、假隔膜及种子微形态系统分类研究的相关报道较少[13~15]。由于成熟的果实及种子具有典型的形态特征,且在同属内、组间具有一定相似性[16]。同时,受遗传因素控制,性状相对稳定,受生态环境影响较小[17~18]。以往对于十字花科果实研究多数集中于开裂、多态性、生殖分析等;种子研究多针对种皮表面纹饰特征、种子的化学组分、萌发、贮藏等[19~21]。本文通过对独行菜族代表植物8属25种果皮的细胞结构、假隔膜结构、种皮细胞结构、子叶与胚根的位置关系进行形态特征比较分析,同时以菥蓂属(Thlaspi)菥蓂(T.arvense)为例详细说明假隔膜的形成过程,为进一步深入研究独行菜族提供更充分详实的形态学依据。
1 材料和方法
1.1 实验材料
1.2 实验方法
取材和固定:采集新鲜种子清洗后放入FAA(Formalin-acetic acid-alcohol mixture)固定液抽气固定(时间不少于24 h);标本保存下的风干种子需先进行复水,待其植株果实及种子基本吸水膨胀后进行固定。
脱水和渗透:根据Feder和O’Brien[22]的方法,将材料先后进行50%酒精、100%酒精、异丙醇及正丁醇脱水(每步需要反复2次,每次不少于4~6 h)、GMA(乙二醇甲基丙烯酸酯)渗透,前2次均不少于24 h,第3次不少于5 d。
包埋和切片:将材料浸入GMA中用胶囊包埋,置于60℃温箱24 h。用Leica Ultralcut R切片,厚度为1.5~2 μm。切片在蒸馏水中展开,置于加热板上烘干。
染色、封片、照相:将切片用希夫试剂—考马斯亮蓝染色,中性树胶封片。在Olympus BX51显微镜下观察,并用Olympus DP70成像系统照相。
果实及种子形态术语参考Vaughan J G、安争西、刘长江等[23~26]的形态描述。
2 结果与分析
2.1 果实及种子微形态结构
荠属(Capsella):短角果,果实呈倒三角形,长5~8 mm,宽4~7 mm,无翅,两侧压扁,无毛,顶端微凹,裂瓣具网脉。外果皮细胞壁较厚,大小不均;中果皮1层纤维层,无木化细胞,维管束为8~10(每心皮),不发达;内果皮为1层较小的厚壁细胞,偶有木化。假隔膜为2层薄壁细胞,如荠(图1:A)。
种子2行,长1 mm,椭圆形浅褐色,单层网纹,无翅,遇水或潮湿环境可形成粘液种子。种皮为3~4层薄壁细胞,外侧细胞较大,形状不规则,内侧细胞均匀有内含物,子叶背倚,如荠(图2:H)。
运转不透明,职责划分不清,没有建立起一套行之有效的监督和问责机制。一是审计难以全覆盖。专项转移支付项目庞杂,即使是大额的专项转移支付,分配到市县后,每个项目规模也不大,难以进行全面审计,更不能形成常态化、普及化的监督规模。二是监管职能分散。目前审计、财政、主管部门对专项转移支付都负有一定的监管职责,但职能重叠,加之部门间信息封闭,监管能力难以整合形成合力,部分专项转移支付存在监管真空。三是监管手段单一。目前对专项转移支付的监管主要是事后检查,许多是问题暴露以后才组织检查,缺乏事前和事中的有效监管。
群心菜属(Cardaria):短角果,卵形或近球形,长3~4.5 mm,宽3.5~5 mm,无翅,两侧压扁,无毛,有明显网脉。外果皮细胞壁较厚,大小不均;中果皮为多层堆叠的薄壁细胞和1层纤维层,无木化细胞,维管束较多,为18~20(每心皮)很发达;内果皮为1层薄壁细胞。假隔膜为2层薄壁细胞,如群心菜(图1:B)。
种子1个,长2 mm,卵形,棕色,单层网纹,无翅,遇水或潮湿环境可形成粘液种子。种皮由4~6层细胞构成,外侧细胞呈波浪形,向内有1层纤维,无木化细胞,内侧细胞有内含物,子叶背倚,如毛果群心菜(图2:G)。

表1 用于研究果实及种子结构的独行菜族物种
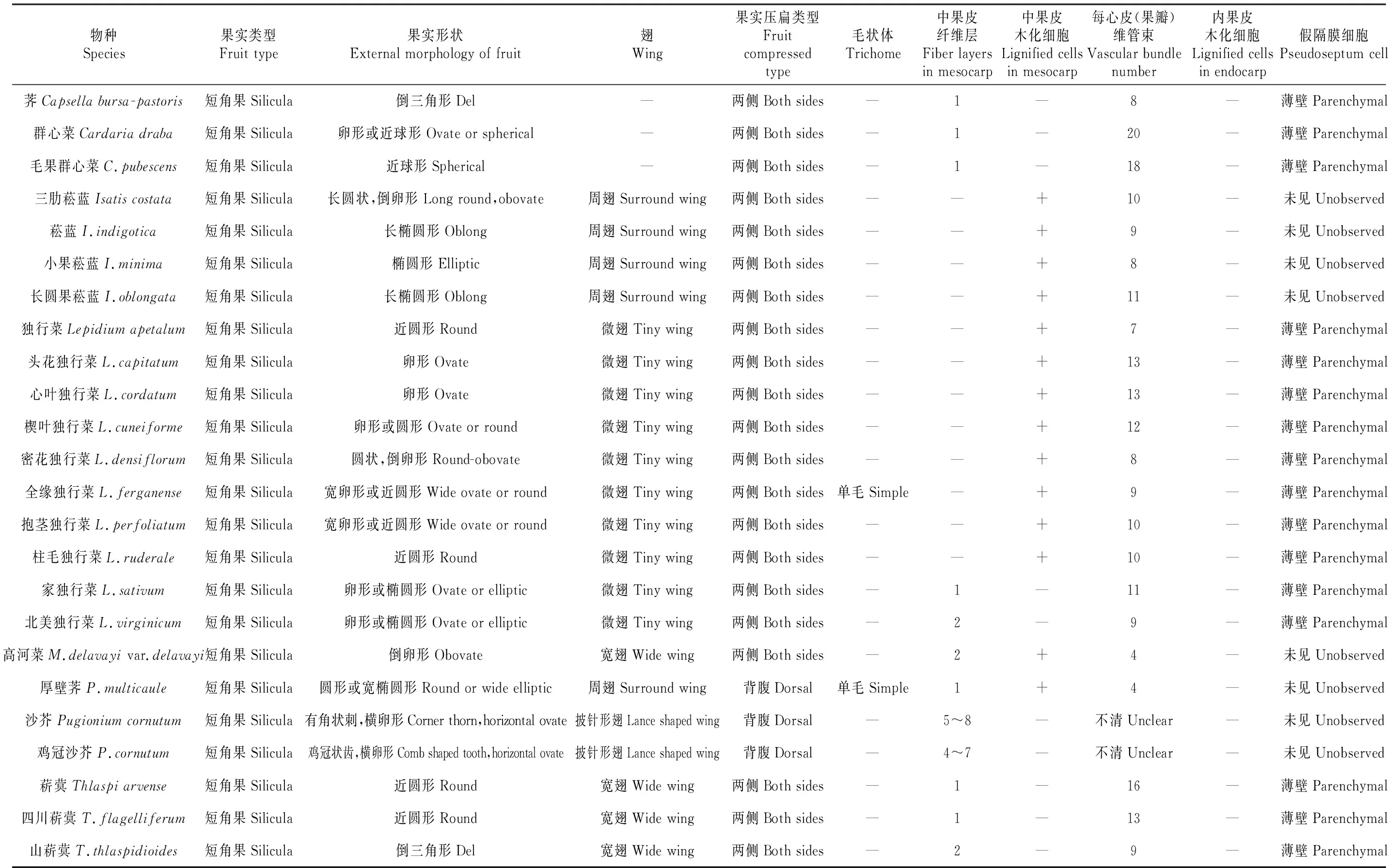
表2 独行菜族果实的外形及结构
“—”.Absent;“+”.Present

图2 果皮及种皮细胞结构 A.山菥蓂翅边缘果皮及种皮细胞结构;B.厚壁荠果皮及种皮细胞结构;C.高河菜果皮细胞结构;D.独行菜果皮及种皮细胞结构;E.长圆果菘蓝果实横切面细胞结构;F.沙芥果皮及刺的细胞结构;G.毛果群心菜果皮及种皮细胞结构;H.荠果皮及种皮细胞结构Fig.2 Cell struture of pericarp and seed epidermis A.T.thlaspidioides; B.P.multicaule; C.M.delavayi var.delavayi; D.L.apetalum; E.I.oblongata; F.P.cornutum; G.C.pubescens; H.C.bursa-pastoris

图3 假隔膜的形成过程 A.前期;B.中期;C.后期;D.末期Fig.3 Forming process of pseudoseptum A.Earlier; B.Medium; C.Anaphase; D.Ttelophase
菘蓝属(Isatis):短角果,长椭圆形,稍扁,长8~15 mm,宽1~4 mm,周翅,两侧压扁,无毛,顶端平截或尖凹,果瓣常有1显明中脉。外果皮为1层薄壁细胞,较小;中果皮细胞木化,大小不均匀,中间偏大两侧偏小,维管束分布在果皮边缘,为8~10(每心皮),向内有1层纤维层;内果皮为1层薄壁细胞,未见假隔膜,如三肋菘蓝(图1:C)。
种子1个,长2~3.5 mm,长椭圆形,褐色,单层网纹,无翅,遇水不形成粘液膜,种皮由多层细胞构成,细胞木化,有内含物,子叶背倚,如长圆果菘蓝(图2:E)。
独行菜属(Lepidium):短角果,近圆形或卵形,略扁,顶端微缺,长2~6 mm,宽约2 mm,两侧有微翅,有毛或无毛,两侧压扁,果瓣偶有龙骨状突起。外果皮为1层椭圆形薄壁细胞构成,大小相近,具角质层;中果皮为多层薄壁细胞紧密排列,无木化细胞,维管束7~10(每心皮)不发达,腹缝线处多为明显的木化细胞积聚,向内有1层纤维层;内果皮为1层薄壁细胞。假隔膜为2层薄壁细胞,如抱茎独行菜(图1:D)。
种子2个,长1 mm,椭圆形或卵形,浅褐色或棕红色,疣状突起或单侧网纹,无翅或有翅,遇水或潮湿环境形成粘液种子。种皮为多层细胞,外侧细胞呈方形,较大,有明显的内含物,向内有3~5层薄壁细胞,偶有木化,子叶背倚,如独行菜(图2:D)。
高河菜属(Megacarpaea):短角果,两果瓣压扁,较大,横8形,顶端宽深陷,长10~14 mm,宽7~10 mm,周翅,两侧压扁,有毛。外果皮由多层薄壁细胞堆叠;中果皮细胞木化,两侧位于翅的位置木化细胞较多,中部偶有单层木化细胞,维管束约2~4(每心皮);内果皮1层薄壁细胞。假隔膜为2层薄壁细胞,如高河菜(图1:E,图2:C)。
种子1个,长5 mm,卵形,棕色,单层网纹,窄翅,遇水不形成粘液膜。种皮为多层细胞,内侧木化,有内含物,子叶缘倚。
厚壁荠属(Pachypterygium):短角果,圆形或卵形,棕色,长2~4.5 mm,宽1~2.5 mm,周翅,背腹压扁,单毛,有不明显中脉。外果皮为多层薄壁细胞叠加;中果皮细胞形状不规则,较大,木化,维管束靠近边缘约4~6(每心皮),较发达;内果皮为1层薄壁细胞。未见假隔膜,如厚壁荠(图1:F)。
种子椭圆形,长1.5 mm,深褐色,双层网纹,窄翅,遇水不形成粘液膜,种皮1~2层薄壁细胞,偶有木化,有内含物,子叶背倚,如厚壁荠(图2:B)。
沙芥属(Pugionium):短角果,革质、角状刺横卵形,长13~16 mm,宽7~13 mm,披针形翅在两侧,中部有角状刺,背腹压扁,无毛,具明显凸起网脉。外果皮为2~3层薄壁细胞;中果皮细胞木化仅在凸起网脉处,维管束不易观察到。内果皮无木化细胞,未见假隔膜,如沙芥(图1:G,图2:F)。
种子1个,长10 mm,长圆形,棕色,较大,横向生长,遇水或潮湿的环境形成粘液种子。多层细胞内侧木化,有内含物,子叶背倚。
菥蓂属(Thlaspi):短角果,近心形,膜质,顶端凹入,长7~10 mm,宽2~4 mm,宽翅,两侧压扁,无毛,无脉。外果皮为1层薄壁细胞,形状较规则;中果皮为3~5层薄壁细胞堆叠,向内有1层纤维层,靠近两侧翅的位置各有1个较大的维管束;内果皮1层薄壁细胞,假隔膜为2层薄壁细胞,如菥蓂(图1:H~I)。
种子4~8个,长1.5 mm,倒卵形,褐色,棱纹或单层网纹,窄翅,遇水或潮湿环境形成粘液种子。种皮有多层薄壁细胞无木化,有内含物,子叶缘倚,如山菥蓂(图2:A)。
2.2 假隔膜的形成过程
独行菜族果实为短角果,开裂或不开裂,2心皮合生,侧膜胎座。根据假隔膜形成过程按照前、中、后、末4个时期进行半薄切片观察。前期:子房中部内表皮细胞逐渐向内腔生长,出现空腔(图3:A);中期:子房生长中空腔逐渐扩大,中部腹缝线处突出的分生组织开始相对生长,较多的薄壁细胞向中间延伸(图3:B);后期:紧密排列的薄壁细胞向内逐渐变得疏松,向内相对生长的薄壁细胞变成2列(图3:C);末期:薄壁细胞相互延伸连接形成一个整体,最终发育成成熟的假隔膜革质,将子房分为2室,胚珠分别着生在假隔膜的两侧(图3:D)。
3 讨论
在我国独行菜族20属中,包括荠属(1种)、群心菜属(3种)、菘蓝属(6种)、独行菜属(15种)、高河菜属(3种)、厚壁荠属(1种)、沙芥属(4种)、菥蓂属(6种)、岩荠属Cochlearia(13种)、臭荠属Coronopus(2种)、双脊荠属Dilophia(3种)、蛇头荠属Dipoma(2种)、藏荠属Hedinia(1种)、半脊荠属Hemilophia(2种)、薄果荠属Hymenolobus(1种)、屈曲花属Iberis(2种)、弯梗芥属Lignariella(1种)、双果荠属Megadenia(1种)、宽框荠属Platycraspedum(1种)、革叶荠属Stroganowia(1种)。大部分属下包含种数目较少,其中14个属内仅包含1~3种植物,12属中6属均为单属种植物,且分布于我国西南部高山或喜马拉雅山等海拔较高的生境下,故未能收集到。在本研究中8个典型代表属的种类所采样品数量均超过本属种数的一半且包含属的模式种,可以为本研究提供相关依据。
本研究中独行菜族果实均为短角果,性状丰富多样,具有重要分类价值。除荠属、群心菜属外果实均具翅可分为4种类型:微翅、宽翅、周翅、披针形翅。其中,厚壁荠属和沙芥属果实为背腹压扁,其他均为两侧压扁;荠属、独行菜属、菥蓂属果实开裂,其他属果实不开裂。果实通常无毛,偶有柔毛,除菥蓂属外,其他属常具网状脉、中脉或龙骨突起。除菘蓝属、厚壁荠属、沙芥属无假隔膜结构外,其他5属均具有明显的假隔膜。部分果皮细胞有纤维层,偶有木化,木化细胞通常在具翅的果实中较为明显,维管束发达。
种子较小,大小为(0.8~10)mm×(0.5~2.8)mm,种皮纹饰丰富,通常遇水或潮湿环境可形成粘液种子,种皮通常由薄壁细胞构成,偶有木化,常存在有内含物。除高河菜属和菥蓂属子叶与胚根的位置关系为缘倚外,其他属均为背倚。
独行菜属为独行菜族代表属种,我国有15种,本研究中包括该属10个物种,前人研究将其分为具瓣组、厚翅组、少蕊组,该属果皮无毛,具翅类型丰富,果皮细胞结构中仅家独行菜中果皮无纤维层、无木化细胞,支持将其与其他种分开,划分为具瓣组[1]。厚翅组仅有1外来物种(Lepidiumcampestre),少蕊组分类性状较稳定。
种皮表面纹饰分为四种类型:荠属、群心菜属、菘蓝属为单层网纹,独行菜属为疣状突起和单层网纹,厚壁荠为双层网纹,菥蓂为棱纹和单层网纹。部分独行菜属、高和菜属、厚壁荠属、菥蓂属具翅,均为窄翅。除菘蓝属外其他属种子遇水均形成粘液种子,属间种皮纹饰性状较稳定,支持前人研究将种皮纹饰作为十字花科分类、鉴定依据[18~20]。
支持孙稚颖根据分子系统学聚类分析将菘蓝属应移出独行菜族,本研究中菘蓝属与独行菜族其他类群的种子不同,遇水或潮湿的环境不形成粘液种子。前人分析菥蓂属和菘蓝属、群心菜属和独行菜属有较近的亲缘关系,本研究中未见相关支持证据[2]。独行菜族中菘蓝属、厚壁荠属、沙芥属无假隔膜且厚壁荠属和沙芥属与独行菜族其他属果实压扁方式不同,二者的亲缘关系及系统位置研究需要其他相关证据。前人研究认为假隔膜不可作为分类依据,与果实开裂有一定联系,本研究中并未发现相关证据,需要进一步探讨[27]。
