烟草合子时期表达基因NtZE1的克隆及结构分析
2019-07-22左紫怡
罗 岸,左紫怡,焦 雄,刘 夏
(长江大学生命科学学院,湖北 荆州 434023)
合子是被子植物生长发育的起点,对其研究具有重要的科学意义。烟草Nicotiana tabacum合子和胚胎细胞的发育模式较为规范,一直被当作研究被子植物胚胎发生的模式植物。在烟草中,精卵结合形成合子后会经过一段时间的休眠,紧接着发生一系列形态变化,比如体积相比卵细胞有所缩小、大液泡逐渐消失、细胞核移动到细胞中央等。之后随着合子完成极性伸长变成长形合子,细胞核移动到合子的合点端,合子进入分裂周期并启动后续的胚胎发育[1]。以上过程伴随着一系列精确的分子调控,其中仍有很多问题不甚清楚。
作者在前期工作中制作了烟草长形合子和卵细胞的cDNA 文库,并且通过差减分析获得大量在合子中特异表达的基因。本研究以其中一个新型基因NtZE1的EST 序列为基础,获得其完整的编码序列(Coding Sequence,CDS)和5’侧翼序列(启动子和5’UTR),并对该基因编码的蛋白质基本理化性质进行分析,对其5’侧翼序列启动基因表达的活性进行验证,为后续基因功能分析等研究奠定基础。
1 材料与方法
1.1 材料
1.1.1菌株与质粒大肠杆菌感受态细胞DH5α购自武汉转导生物科技发展有限公司,实验所需质粒由武汉大学彭雄波副教授赠予。
1.1.2植物材料烟草Nicotiana tabacum红花品种SR1、小叶烟草N.benthamiana种植于长江大学生命科学学院温室中,温室条件为恒温25 ℃,光暗周期16 h/8 h。
1.1.3主要试剂EasyPure Quick Gel Extraction Kit试剂盒、EasyPure Plasmid MiniPrep Kit试剂盒、EasyPure PCR Purification Kit试剂盒、限制性内切酶购自北京全式金生物技术有限公司;Taq酶、Ex Taq酶、PrimeSTAR@GXL高保真酶、T4 DNA Ligase购自TaKaRa公司;Universal Genome Walker Kit试剂盒购自Clontech公司;Dynabeads®mRNA DIRECT™ Micro Kit试剂盒购自赛默飞世尔科技(中国)有限公司;SMART-Seq v4 Ultra Low Input RNA Kit for Sequencing购自Clontech公司。
1.2 方法
1.2.1烟草基因组文库获得和NtZE1基因5’侧翼区克隆取烟草幼叶按照CTAB 法制得基因组,将其按照Universal Genome Walker Kit 试剂盒的要求进行处理,获得合格的基因组文库。用于克隆的GSP引物、反应条件和体系按照试剂盒的要求设置。扩增得到的目的片段测序后进行序列拼接和延伸。全长检测引物Pro-F:TGGGTTGATACTTGATGGTTTTGG;Pro-R:GGTTGTATTTTCTTTTTCCTTTTTC TATT。
1.2.2烟草长形合子cDNA的获得和NtZE1基因编码序列验证应用传统的酶解-研磨法分离烟草长形合子[2—3],收集约50个合子用于后续操作。按照Dynabeads® mRNA DIRECT™ Micro Kit试剂盒的说明分离出长形合子mRNA,并使用SMART-Seq v4 Ultra Low Input RNA Kit for Sequencing试剂盒制成长形合子cDNA。以烟草长形合子cDNA为模板,用引物CDS-F1/CDS-R1扩增NtZE1基因的CDS序列,CDS-F:ATGAATCTCAAAAGGGTGTTTGTAT;CDS-R:TTAATTTCCAAGTTTTTTCGACTTGT。
1.2.3NtZE1基因启动子的EGFP核定位表达载体构建根据序列信息设计NtZE1基因5’侧翼区扩增引物,并在引物5’端添加合适的酶切位点用于基因克隆。以烟草基因组为模板扩增NtZE1基因的5’侧翼序列(启动子和5’UTR)。扩增产物胶回收后进行双酶切,相应的酶切产物纯化后连接到核定位表达载体。
1.2.4NtZE1基因启动子活性检测构建NtZE1基因启动子的EGFP 核定位表达载体。利用瞬时表达技术将上述载体注入小叶烟草叶表皮细胞,48 h 后在荧光显微镜下观察。
1.2.5生物信息学分析用Omega 软件分析基因序列中的开放阅读框;用ProtParam、ProtScale 等在线软件分析蛋白质一级结构(包括疏水性和不稳定性等理化性质);用SOPMA、SWISS-MODEL 等在线软件分析蛋白质二、三级结构;用PSORT、BaCelLo、SignalP 等在线软件分析蛋白质信号肽的存在和蛋白质在亚细胞中的定位;用PLACE 和PlantCARE 等在线软件分析启动子序列。
2 结果与分析
2.1 烟草合子的分离
图1 显示,利用酶解和显微操作分离得到的处于不同发育阶段的合子大小形状和生活状态均属正常。因此,按此方法可保证收集到数量足够、质量合格、时期准确的长形合子,用于制备其cDNA 作为基因克隆的模板。
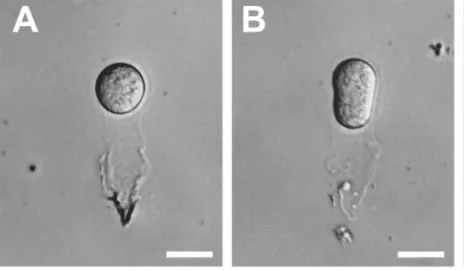
图1 烟草不同发育阶段的合子的分离Fig.1 Isolation of zygote from different developmental stages in tobacco
2.2 烟草NtZE1 基因的克隆
通过差减分析前期获得的烟草长形合子和卵细胞的cDNA 文库,获得长度为600 bp 的NtZE1基因EST 序列。经Omega 软件分析发现其具有一个315 bp 开放阅读框(图2: A)。以重新收集的长形合子细胞制作的cDNA 为模板,可以扩增到该基因的CDS 序列,证实NtZE1基因参与了合子时期的发育事件(图2: B)。此外,利用NCBI 数据分析NtZE1基因的CDS 序列和编码氨基酸序列,发现除在绒毛状烟草N.tomentosiformis中有一条较为类似的序列外,尚未在其他物种中发现类似序列,说明NtZE1基因可能是在合子生长发育中行使未知功能的新型基因。利用已知的EST 序列为基础,运用Genome walking技术进一步获取NtZE1基因5’侧翼序列2578 bp(包括启动子区和5’UTR 区)(图2: B)。
2.3 NtZE1 蛋白基本理化性质
使用ProtParam 在线软件对NtZE1蛋白进行分析。结果表明,该蛋白由105 个氨基酸残基组成,分子量为12.0522 kDa。在105 个氨基酸残基中共包含19 种基本氨基酸类型,但缺少色氨酸,且在所有氨基酸中半胱氨酸和赖氨酸的比例最高,各占10.5%。针对氨基酸残基性质的分析表明,NtZE1蛋白拥有14 个正电荷残基和11 个负电荷残基。NtZE1蛋白的理论等电点为8.28,属于碱性蛋白。该蛋白的脂肪族氨基酸指数为70.57,不稳定性指数为49.10,平均总疏水性为-0.023。因此,NtZE1蛋白应为亲水性蛋白,且不太稳定(表1)。
2.4 NtZE1 蛋白二、三级结构预测
用SOPMA 在线软件对NtZE1蛋白二级结构预测,显示α-螺旋占69.52%,延伸链占3.81%,β-转角占3.81%,无规则卷曲占22.86%(表2)。图3 显示,α-螺旋在NtZE1蛋白中含量最多,分布最为广泛,而无规则卷曲主要分布在蛋白中部和C 端。这与使用SWISS-MODEL 服务器预测的蛋白质三级结构基本吻合(图4)。

图2 烟草NtZE1 基因的克隆Fig.2 Cloning of NtZE1 in tobacco

表1 NtZE1 蛋白基本理化性质的分析Table 1 Basic physicochemical properties analysis of NtZE1 protein
2.5 NtZE1 蛋白亚细胞定位及信号肽序列
利用ProtScale 在线软件分析,发现位于NtZE1蛋白N 端的氨基酸可能具有较高的疏水性。进一步使用SignalP 软件分析,发现在NtZE1蛋白N端存在信号肽,且其剪切位点位于第23 和24 个氨基酸之间(图5)。这与预测的NtZE1蛋白的亚细胞定位较为一致,PSORT、BaCelLo 等在线软件预测,NtZE1蛋白在细胞外分布的可能性最大。

表2 NtZE1 蛋白二级结构的预测Table 2 Prediction of NtZE1 protein secondary structure

图3 NtZE1 蛋白二级结构Fig.3 Prediction of NtZE1 protein secondary structure

图4 NtZE1 蛋白三级结构Fig.4 Prediction of NtZE1 protein tertiary structure
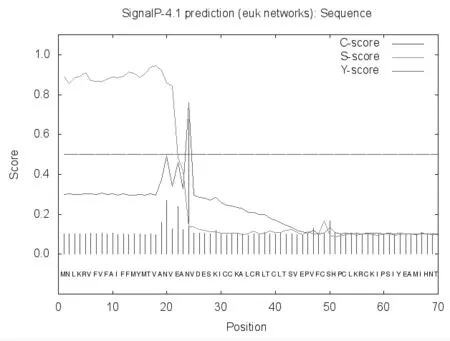
图5 NtZE1 蛋白的信号肽预测Fig.5 Prediction of NtZE1 protein signal peptide
2.6 NtZE1 基因5’侧翼序列分析及其启动基因表达的活性检测
以已知的NtZE1基因EST 序列为基础,运用Genome walking 技术对该序列未知的5’侧翼序列进行延长,最终获得自ATG 起始密码子前共2578 bp 的5’侧翼序列(启动子和5’UTR)。PLACE 和PlantCARE 在线软件预测表明,NtZE1基因的表达调控序列中含有其他大多数基因表达调控序列所含有的TATA-box 和CAAT-box 等基本转录元件。此外,NtZE1基因的5’侧翼序列中可能存在一些与激素、光、胁迫等相关的顺式调控元件,暗示NtZE1基因可能受多种外界信号调控。
为验证所获得的5’侧翼序列(启动子和5’UTR)具有启动基因表达的能力,对序列进行扩增并通过酶切连接到EGFP 核定位载体中,再借助瞬时表达技术将载体注入小叶烟草叶表皮细胞。利用荧光显微镜观察经注射的叶表皮细胞,发现在一些细胞的细胞核中有非常清晰的绿色荧光(图6)。这显示获得的NtZE1基因5’侧翼序列(启动子和5’UTR)启动基因表达的活性较强,能被运用于基因功能分析。

图6 NtZE1 基因5’侧翼序列(启动子和5’ UTR)启动基因表达的活性Fig.6 Gene expression activity of NtZE1 5'flanking sequence ( promoter and 5’ UTR)
3 结论与讨论
NtZE1基因在烟草合子中表达,编码的新型蛋白由105 个氨基酸残基组成。在NCBI 数据库中无法查询到类似的基因序列,因此NtZE1基因的功能暂时未知,其对合子的生长发育可能有独特的贡献。软件分析预测在NtZE1蛋白的N 端存在信号肽序列,该蛋白的亚细胞定位可能在细胞外。
目前已经将发现的若干与合子败育相关的突变体用于对合子的研究。比如在fac1 突变体中合子的发育停滞,原因是在合子中表达的FAC1 基因功能缺失,造成其编码的AMP 脱氨酶(AMPD)无法正常行使功能,因而不能产生更多的能量供给细胞生长所需[4—5]。在gcd1 突变体中合子的发育也出现停滞,原因是线粒体定位的GCD1 蛋白出现异常,造成卵细胞和中央细胞无法成熟,从而影响后续的胚胎发生[6]。同样定位于线粒体中的MIRO1 蛋白如果发生突变亦会影响合子发育,产生合子致死的表型[7]。此外,合子的不等分裂也是植物胚胎发生中的重要事件,对后续胚胎发生过程中的细胞命运决定和胚胎模式建成等具有重要意义。诸如GNOM、RSH、YODA、SSP、GRD、WRKY2、YAO等基因都与合子不等分裂有关[8—17]。不过上述调控因子一般都停留在细胞内发挥作用,而胞外微环境中的信号分子通过何种途径影响合子发育,到目前为止所知甚少。二十多年前,对植物体细胞胚胎发生的初步研究曾提示,外界的化学或物理环境可能影响胚柄的正确形成[18]。近年来对拟南芥Arabidopsis thaliana和玉米Zea mays等的研究也证实,胚胎的正常发育的确会受到外界环境的影响[19—21],但到目前为止,参与调控植物合子发育的胞外信号到底有哪些,其具体的分子机理又是什么,所知仍近空白。NtZE1基因的发现提示一种新型未知功能的胞外蛋白可能在烟草合子的生长发育中发挥着重要作用。本研究以先期获得的NtZE1基因的EST 序列为基础,获得了其5’侧翼序列(启动子和5’UTR)。对5’侧翼序列进行克隆后连入EGFP 核定位载体并在小叶烟草叶表皮细胞中瞬时表达,结果显示在细胞核中能观察到明显的绿色荧光。这说明本研究所获得的NtZE1基因的5’侧翼序列(启动子和5’UTR)具有较强的启动基因表达活性,为以后分析NtZE1基因的表达模式、基因功能等奠定了基础,有助于丰富对合子发育调控机制的认识。
