猪脂肪沉积相关的MicroRNA研究进展
2019-07-22陈辉
陈 辉
(华中农业大学农业动物遗传育种与繁殖教育部重点实验室,农业农村部猪遗传育种重点开放实验室,湖北 武汉 430070)
猪肉是国人主要的食用肉类,而随着人民生活水平的提高,对优质猪肉的需求也随之增加。由于前期人们对瘦肉型猪的高强度选择,使得猪肉的风味和多汁性等品质下降[1],而脂肪沉积是影响猪肉品质(风味和多汁性等)的主要因素。MicroRNA(miRNA)最早是在1993年由Lee等在秀丽隐杆线虫中发现的,研究发现了一种可以与Line4基因的3'非翻译区(3'UTR)形成互补序列从而抑制该基因的翻译,这是MicroRNA首次被发现的标志[2]。miRNA是长度约为 22 nt(18~24nt)的内源性小分子RNA,在动植物机体通过靶向目的基因致使基因表达受阻,从而实现对动植物机体的各项生命活动(脂肪细胞增殖分化等过程)进行调控[3]。有关miRNA的相关研究主要集中在人上,笔者通 过 MicroRNAs(miRNA) 数 据库(http://www.mirbase.org/) 检 索了人(Human)相关miRNA,截止到目前,已有2 815条miRNA数据;笔者同样检索了猪(swine)相关miRNA,结果仅为219条相关miRNA数据,而与猪脂肪沉积相关的miRNA数据就更少了,因而需要进一步深入挖掘和鉴定猪脂肪沉积相关的miRNA。研究表明,miR-27[4]、miR-34[5]、miR-125[6]以及miR-130[7]等miRNA家族可通过靶向目的基因实现对猪脂肪沉积进行调控,故就4个miRNA在猪脂肪沉积中的调控进行综述,同时分析了miRNA的形成及作用机制,旨在为进一步深入挖掘和鉴定与猪脂肪沉积相关miRNA提供理论依据和思路,进而为猪肉品质的遗传改良提供策略和方向。
1 miRNA的形成及其作用机制
1.1 miRNA的形成过程
猪miRNA是在细胞核经RNA聚合酶Ⅱ(RNA Pol II)的转录产物[8]。具体形成过程分为3步。第一步是miRNA基因通过RNA Pol II的转录形成初级具有发卡结构的转录物(pri-miRNA);第二步是通过RNase核酸内切酶III(DROSHA)将pri-miRNA加工形成前体miRNA(pre-miRNA),产生的pre-miRNA(具有二级茎环结构)通过核蛋白(Exportin-5,一种GTP依赖性转运蛋白)转运至细胞质;最后一步即是通过Dicer酶(一种RNA聚合酶III)和TRBP(RNA结合蛋白)和PACT进行切割形成双链miRNA,至此成熟的miRNA加工形成。其具体形成过程见图1[9]。
1.2 miRNA的作用机制
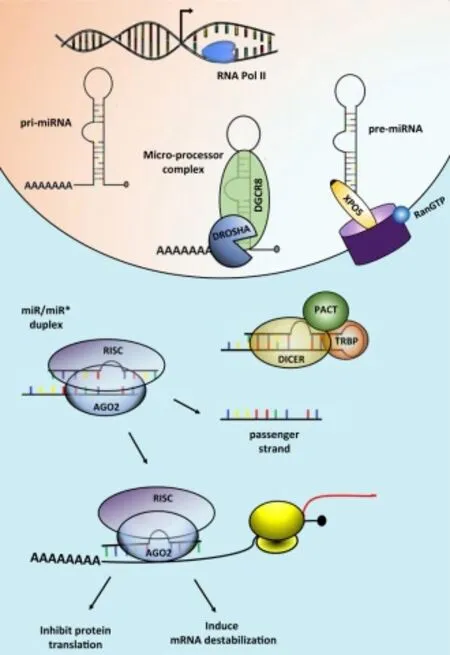
图1 miRNA的形成过程
miRNA主要是形成沉默复合物(RISC)后,用于种子序列靶向目的基因从而抑制基因的表达。在细胞核加工形成双链miRNA,此时的miRNA可以形成活性RNA诱导形成沉默复合物(RISC),RISC中包含的Argonaute 2蛋白(Ago2)是RISC的关键催化酶,一旦Ago2与RISC结合,目的基因的3'UTR将会被miRNA的5'末端的互补序列(称为种子序列,由于种子序列多为6~8 nt,因而一种miRNA可靶向多种基因,同理基因也可被多种miRNA靶向)识别并结合,从而阻止其翻译过程,使靶基因mRNA沉默[8]。
2 猪脂肪沉积相关miRNA
2.1 miR-27家族
miR-27家族包括miR-27a、miR-27b以及miR-27c,研究发现miR-27家族通过靶向脂蛋白酶(LPL)基因,对LPL的表达进行抑制,从而降低了脂肪沉积相关基因的表达,最终抑制了脂肪的沉积,对猪肉品质的改善具有消极作用[10]。研究表明,miR-27在脂肪组织的表达量显著降低(P<0.05),猪皮下脂肪中miR-27的表达与脂肪沉积标志基因过氧化物酶增殖物激活受体(PPARγ)-γ以及脂肪酸结合蛋白4(FABP4)的表达呈显著负相关(P<0.05),这表明miR-27对脂肪沉积具有抑制作用[11]。关于miR-27a的研究,有研究发现miR-27a在藏猪背最长肌的表达量极显著高于在大白猪中的表达量(P<0.01),相关性分析表明,miR-27a的表达量与IMF含量呈负相关关系[12]。相似的研究报道,miR-27a在猪背部皮下脂肪的表达水平与背膘厚以及单不饱和脂肪酸呈负相关关系,而与胴体瘦肉率呈显著正相关,表明miR-27a对猪脂肪沉积具有抑制作用,而有助于改善猪胴体性能[13]。但也有关于miR-27a正调控脂肪沉积的研究,Li等[4]研究发现,在沂蒙黑猪中,背膘较厚的个体中miR-27a具有更高的表达量;杨雪梅等[14]采用荧光定量PCR技术(Q-PCR)探讨了miR-27a的表达量与我国地方猪种肉质性状(风味物质)的关系,结果表明miR-27a的表达量与地方猪种肌内脂肪(IMF)含量呈正相关关系,这表明miR-27a对肉质性状具有正调控作用,造成差异的原因可能是品种原因。在miR-27b的研究方面,研究发现miR-27a和miR-27b在皮下脂肪细胞的表达量显著高于肌内脂肪细胞的表达量(P<0.05),进一步分析表明,miR-27b能够靶向PPARγ基因,从而抑制其表达,降低脂肪的沉积[15]。综上表明,miR-27家族通过靶向脂肪沉积相关基因从而对脂肪沉积具有抑制作用,对猪肉品质具有负调控作用。
2.2 miR-34家族
miR-34家族主要包括miR-34a、miR-34b以及miR-34c,在猪上研究较多的为miR-34a和miR-34c,故本文主要论述miR-34a和miR-34c在猪脂肪沉积中作用。有关miR-34a的研究较多,研究发现,miR-34a过表达增加了肝脏中甘油三脂的含量,转录调控分析表明,miR-34a通过靶向猪脂联素受体2基因(AdipoR2)的3'UTR抑制和AMPK信号途径来抑制PPARα信号传导途径而增加脂肪的含量[5]。李秀秀[16]采用猝灭萤火虫荧光素酶方法确定了miR-34a的种子序列与长链酯酰辅酶A合成酶4(ACSL4)基因的靶向关系,细胞增殖分化分析表明,miR-34a靶向ACSL4基因从而抑制其表达,进而促进脂肪细胞的增殖分化,从而使脂肪细胞脂滴含量增加。但也有miR-34a负调控脂肪沉积的研究报道,Sun等[17]研究了miR-34a与人血小板衍生生长因子受体α(PDGFRα,PDGFRα通过激活Erk信号通路促进脂肪形成分化)的靶向关系,发现miR-34a可以通过靶向PDGFRα,抑制其表达,从而抑制脂肪的生产。在miR-34c与脂肪沉积中的研究中,陈粉粉[18]研究miR-34c对猪肌内脂肪细胞增殖分化的影响,研究发现,过表达miR-34c后,发现肌内脂肪细胞分化标志基因PPARγ、FABP4和FASN基因的转录水平(mRNA)被极显著下调,蛋白印迹得到了类似的结果;流式分析表明,过表达miR-34c抑制了肌内脂肪细胞的增殖;最终抑制了猪脂肪的沉积,表明miR-34c对脂肪沉积具有负调控作用。
2.3 miR-125家族
miR-125a是miR-125家族的主要成员,也是一种猪脂肪沉积的抑制因子,通过抑制脂肪细胞的增殖分化而抑制脂肪的沉积。Du等[6]miR-125a可以直接靶向kruppel 3因子(KLF13),从而抑制猪肌内前脂肪细胞的分化,同时研究发现miR-125a的表达量与IMF含量呈负相关;过表达miR-125a降低了总饱和脂肪酸(SFA)含量和单不饱和脂肪酸(MUFA)/ SFA比率。在对鼠的研究表明,miR-125a可能促进3T3-L1前脂肪细胞增殖而通过负调节STAT3抑制3T3-L1前脂肪细胞分化[19]。Ji等[20]研究表明,过表达miR-125a后会显著降低成脂标志基因PPARγ,LPL和aP2的mRNA表达水平;油红O染色分析表明,过表达后miR-125a显著降低了脂质的积累;通过靶向分析,miR-125a可靶向ERRα基因从而抑制猪脂肪细胞的分化。可抑制细胞的增殖分化而实现对脂肪沉积的负调控,Song等[21]发现,miR-125a可靶向转录因子E2F3的3'UTR,从而负调控E2F3的表达,过表达miR-125a后发现C2C12成肌细胞的增殖被抑制,最终降低了脂肪的沉积。相似的研究发现,过表达miR-125a后会导致脂滴降低,成脂标志基因PPARγ和aP2的表达量以及翻译水平均下调,同时过表达miR-125a显著抑制了猪前体脂肪细胞的分化;而抑制miR-125a则得到了相反的结果(促进了脂肪前体细胞的分化)[22]。表明,miR-125a通过靶向成脂标志基因而抑制脂肪细胞的增殖分化,从而对猪脂肪的沉积起负调控作用。
2.4 miR-130家族
miR-130家族也是一种脂肪沉积的抑制因子,该家族主要包括miR-130a和miR-130b两大类。在miR-130a与猪脂肪沉积的研究方面,wei等[7]研究了肌内脂肪(IM)和皮下脂肪(SC)之间PPARγ的表达水平差异,结果发现IM中的PPARγ的mRNA和蛋白质水平均显著低于SC,而IM中的miR-130a的表达水平为SC的2.30倍,因此作者进一步通过双荧光素酶活性方法探讨miR-130a和PPARγ的表达关系,结果表明miR-130a直接靶向PPARγ,抑制其表达,进而造成IM中的脂肪沉积水平低于SC。miR-130a还可通过靶向成脂标志基因而抑制脂肪细胞分化,据李虹仪等[23]报道,将miR-130a类似物(miR-130a mimics)转染至猪皮下脂肪前体细胞后连续诱导8 d,与阴性(乱序序列)对照相比,miR-130a mimics组细胞脂滴减少,甘油三酯(TAG)含量显著降低,脂肪细胞分化标志基因LPL和PPARγ的表达量均显著下调(P<0.05);而加入miR-130a类抑制物后,细胞脂滴和TAG含量增加,这表明miR-130a可靶向多个基因以实现对脂肪细胞分化的调控。miR-130b也可通过靶向PPARγ等基因来抑制脂肪的沉积,Pan等[24]发现miR-130b可以直接靶向PPARγ基因,降低其mRNA水平,从而抑制猪脂肪的沉积。也有研究通过双荧光素酶活性方法测定了miR-130b的表达与PPARγ、C / EBP-β3 等成脂基因的 3,UTR,抑制目的基因的表达,研究结论认为miR-130b对母猪及其仔猪的脂肪沉积起负调控作用[25]。综上表明,miR-130家族通过靶向成脂相关基因,进而抑制脂肪细胞的增殖分化,最终对猪脂肪沉积进行负调控。
2.5 其他脂肪沉积相关miRNA
除了上述与猪脂肪沉积相关的miRNA外,还有miR-17-5p、miR-103、miR-199-5p、miR-425p以 及miR-7134-3p等miRNA参与了猪脂肪沉积的负调控。研究发现过表达miR-17-5p会显著抑制猪前脂肪细胞 NCOA3、PPARγ以及FABP4基因的表达(P<0.05),同时显著降低TAG的含量(P<0.05);而抑制miR-17-5p表达则结果相反,研究认为miR-17-5p通过靶向NCOA3基因来抑制猪脂肪的沉积[26]。Shi等[27]发现miR-199-5p是通过靶向caveolin-1(Cav-1)来调控猪脂肪细胞的分化;Chen等[28]发现miR-425p可以靶向KLF13,下调PPARγ基因的表达进而抑制猪肌内脂肪的形成;Wang等[29]发现MARK4基因是miR-7134-3p的靶基因,通过抑制基因的表达进而抑制阉猪的脂肪沉积。上述miRNA均是脂肪沉积的负调控因子,而研究表明miR-103具有促进猪脂肪沉积的作用,Li等[30]研究表,过表达miR-103后会促进脂肪细胞的分化,增加诱导8 d后的脂滴积累,同时促进成脂标志基因PPARγ基因、aP2基因的表达量,研究认为PPARγ基因有助于猪原代脂肪细胞中的脂肪沉积,从而对猪脂肪沉积起正调控作用。
3 小结与展望
通过全文可知,miRNA通过种子序列靶向目的基因的3,UTR,进而从转录后水平对目的基因的表达进行抑制,实现对基因表达的调控。miR-27、miR-34、miR-125以及miR-130家族参与了猪的脂肪沉积 调 控,miR-27、miR-125、miR-130家族为脂肪沉积的负调控因子,miR-34a以及miR-103则是猪脂肪沉积的促进因子,这丰富了miRNA在猪脂肪沉积中的研究,为猪肉品质的遗传改良提供了新的策略和思路。尽管miRNA在脂肪调控中具有重要调控作用,但有关miRNA在猪脂肪调控方面的研究却鲜有报道;同时大部分miRNA相关研究只是复制人或小鼠的实验结果,并未进行深入系统的研究,部分文章虽然论述了miRNA通过靶向脂肪沉积相关基因,但却缺乏科学合理的实验设计;总之,与miRNA在人和鼠上的研究相比,miRNA调控猪脂肪沉积方面的研究远远不够[31]。但相信随着分子生物技术、猪遗传改良技术的发展,科学家将会对miRNA调控猪脂肪沉积方面的研究进行更加深入系统的研究,同时鉴定出更多的与脂肪沉积相关miRNA,从而丰富miRNA的相关研究,也为猪肉品质的遗传改良提供新的策略和思路。
