东北七鳃鳗早期发育研究
2019-07-20马庆华刘怀秀滕洪明欣韩英伦李庆伟
李 军 马庆华 刘怀秀 宋 涛 滕洪明 朱 婷 刘 欣韩英伦 李庆伟
(1. 辽宁师范大学生命科学学院, 七鳃鳗研究中心, 大连 116081; 2. 辽宁省淡水水产科学研究院, 辽宁省水生动物病害防治重点实验室, 辽阳 111000)
七鳃鳗(Lamprey)是圆口纲无颌类脊椎动物,现存脊椎动物中最古老的类群[1]。由于重要的进化地位, 七鳃鳗已成为研究发育生物学的关键物种[2,3]。七鳃鳗早期发育主要分为胚胎、卵黄囊仔鱼和幼鱼三个发育时期[3]。研究七鳃鳗早期发育可以了解胚胎、卵黄囊期仔鱼和幼鱼发育过程, 为七鳃鳗的发育学研究提供基础资料[4]。目前对七鳃鳗胚胎发育和卵黄囊期仔鱼的生长发育已有一些研究报道, 如Lampetra fluviatilis[5]、海七鳃鳗(Petromyzon marinus)[6]、Entosphenus tridentatus[7]和日本七鳃鳗(Lampetra japonica)[8]等。研究表明, 七鳃鳗胚胎和卵黄囊期仔鱼的发育过程相似; 七鳃鳗胚胎具有与模式动物胚胎(爪蟾和斑马鱼)相似的生物学特性, 是开展发育生物学研究的良好实验材料[9,10]。而七鳃鳗幼鱼的生长发育研究却鲜有报道, 这与七鳃鳗幼鱼不易采捕以及野外样本的不确定性[11]有关。
东北七鳃鳗(Lampetra morii)是我国特有种[12,13]。相对于其他七鳃鳗, 东北七鳃鳗作为发育生物学的动物模型更具优势: (1)易于养殖, 东北七鳃鳗终生栖息于淡水环境[1,13], 易于采捕, 易于进行增养殖研究;(2)个体较小, 东北七鳃鳗成体大小为160—250 mm,对比日本七鳃鳗(300—500 mm)[1,14]和海七鳃鳗(114—1200 mm)[15]等洄游型种类个体较小, 属小型七鳃鳗, 易于实验操作; (3)生长周期短, 东北七鳃鳗生长周期为4—5年[1,13], 相对于日本七鳃鳗(6—7年)[1,13]和海七鳃鳗(5—8年)[16]等生长周期短, 易于实现人工养殖; (4)怀卵量大, 东北七鳃鳗怀卵量约10万粒[13], 可为试验提供充足的胚胎和仔鱼。东北七鳃鳗的生物学特点使其成为七鳃鳗研究中一个优良物种, 而目前尚未有东北七鳃鳗胚胎发育、卵黄囊期仔鱼和幼鱼的生长发育的报道。本研究开展东北七鳃鳗胚胎、卵黄囊期仔鱼和幼鱼发育研究, 较为系统地描述东北七鳃鳗的早期发育形态特征和生长发育规律, 以期为七鳃鳗发育学研究提供基础资料, 推进七鳃鳗的模式化进程。
1 材料与方法
1.1 材料
东北七鳃鳗亲本[雌性: (22.78±2.37) g, (22.76±1.85) cm; 雄性: (27.08±5.03) g, (22.51±1.26) cm]采捕于鸭绿江水丰水库(2015年4月至6月)。
东北七鳃鳗胚胎采用人工授精方法获得, 在繁殖季节, 根据东北七鳃鳗副性征及性腺成熟度, 共选取45尾亲本(雌性15尾, 雄性30尾), 采用人工授精方法获得胚胎[6,9]。当雌雄亲本具有明显的副性征和繁殖行为时, 用0.01% MS-222麻醉, 用手轻轻挤压亲鱼的腹部, 将成熟的卵盛放在烧杯中(盛装约300 mL养殖用水), 立即用同样的方法向烧杯内挤入精液, 混匀, 漂洗3—5次, 即可布入孵化缸, 获得胚胎。
东北七鳃鳗卵黄囊期仔鱼和幼鱼均由胚胎培育获得。
1.2 方法
东北七鳃鳗养殖、孵化和培育管理东北七鳃鳗亲本在养殖系统中暂养, 养殖系统由水族箱(100 cm×50 cm×50 cm)、过滤系统和黑色陶粒砂(2—3 mm3)组成, 陶粒砂铺在水族箱底部(10—15 cm高), 水高约40 cm, 水流约2 L/min。放养密度为30尾东北七鳃鳗/水族箱, 养殖水温为5—18℃(11月至次年5月), 溶解氧≥5.0 mg/L, pH 7.0±0.2。
受精卵、初孵仔鱼和幼鱼均在孵化缸内孵育,孵化缸由水族箱(50 cm×50 cm×50 cm)和养殖用沙粒(约0.5 mm3)组成, 每个孵化缸盛水高约40 cm, 缸底部铺有约1.5 cm厚养殖用沙粒; 孵化缸内每2枚胚胎间保持间距约为1 cm; 孵化缸内投放初孵仔鱼和幼鱼的密度约200尾/缸。受精卵的孵化、初孵仔鱼和幼鱼培育采用静水养殖, 用气泵充气, 胚胎孵化和初孵仔鱼培育水温(18±1)℃, 幼鱼培育水温(19±2)℃, 溶解氧≥5.0 mg/L, pH 7.0±0.2。
培育期间, 室内保持安静、弱光, 每天换等温水1次, 换水量约为孵化缸内水量的1/3, 养殖用水为充分曝气至少3d的自来水。
东北七鳃鳗胚胎发育东北七鳃鳗胚胎发育观察参照日本七鳃鳗胚胎发育观察方法[8]。在体视镜下(SMZ18, Nikon)观察其发育形态特征和记录胚胎发育起始时间, 当其中50%以上胚胎出现某个时期发育特征则记为该发育阶段的起始时间。分别收集各发育阶段胚胎50—100枚, 用4%多聚甲醛固定, 4℃保存, 待东北七鳃鳗胚胎发育结束后, 在解剖镜下观察、照相。
东北七鳃鳗卵黄囊期仔鱼的发育和异速生长
试验选取同一天破膜孵出的仔鱼1000尾用于试验, 待仔鱼卵黄囊完全吸收时实验结束, 实验周期为16d。每天取出30尾仔鱼, 用浓度为0.01% MS 222麻醉, 在体视镜下观察其外部形态特征、照相,应用NIS-Elements D 4.60.00软件测量吻长、眼径、眼鳃间距、口笠长、鳃前长、鳃长、头长、体长、尾长、泄殖孔长和全长各形态学指标(精确至1 μm, 图 1); 体重用分析天平测量, 每次称量10尾仔鱼(精确至0.0001 g), 共取3次, 取平均值。
东北七鳃鳗幼鱼发育试验选取同一天破膜孵出500尾仔鱼用于试验。每月取出30尾幼鱼,幼鱼用浓度为0.01% MS 222麻醉, 在体视镜下观察其外部形态特征、照相, 应用软件(NIS-Elements D 4.60.00软件)和游标卡尺测量全长(精确至1 mm,图 1), 用分析天平测量仔鱼体重(精确至0.0001 g)。
1.3 数据统计与分析

图 1 东北七鳃鳗卵黄囊期仔鱼测量图Fig. 1 Illustration of measurement of Lampetra morii yolk-sac larva1. 吻长; 2. 眼径; 3. 眼鳃间距; 4. 口笠长; 5. 鳃前长; 6. 鳃长; 7.头长; 8. 体长; 9. 尾长; 10. 泄殖孔长; 11. 全长1. snout length, 2. eye length, 3. postocular length, 4. disc length,5. prebranchial length, 6. branchial length, 7. head length, 8. trunk length, 9. tail length, 10. cloacal slit length, 11. total length
2 结果
2.1 东北七鳃鳗亲本形态性状和繁殖行为
在非繁殖季节(9月至次年2月), 东北七鳃鳗成体无副性征, 不易鉴别雌雄。大多数时间, 东北七鳃鳗聚集在水族箱的一角, 极少游动。在水族箱中投放饵料鱼时, 东北七鳃鳗逐渐活跃、靠近、叮咬饵料鱼, 开始摄食。在繁殖季节(4—6月), 东北七鳃鳗十分活跃, 其在水族箱内来回游动, 并不时向水面窜跃, 部分亲本用口吸盘吸住缸壁, 或用口吸盘吸住较大石块, 成排排列, 时而剧烈摆动尾部, 搅动底层陶粒砂。东北七鳃鳗在繁殖期间并未见其摄食活动。
在繁殖季节, 性成熟东北七鳃鳗具有副性征(图 2A、2B), 随成熟度增加, 背部第一背鳍和第二背鳍相互靠近, 直至2个背鳍在基部相连, 矗立在背部上方, 似山峰状(图 2A-a、2A-b、2B-a、2B-b);背鳍基部膨大(图 2A-b、2B-b); 背鳍基部和中部充血, 可见毛细血管(图 2A-b、2B-b); 泄殖孔肿胀(图2A-c、2B-c)。东北七鳃鳗雌雄性的副性征并不完全相同, 雌性头后至泄殖孔间腹部膨大, 性腺的外形明显(图 2A-a); 第二背鳍基部凸起(图 2A-a、2A-b); 尾部明显上翘(图 2A-a); 臀鳍增厚膨大(图 2A-b、2A-c); 泄殖孔表现出肿胀充血(图 2A-b、2A-c),生殖孔凸出于泄殖孔(图 2A-c), 解剖可见腹腔内乳白色卵巢, 卵圆型卵游离于卵巢中。而雄性则表现出头部和尾部稍向下弯, 其腹部稍膨大, 明显小于雌性(图 2B-a); 细长的生殖孔凸出于泄殖孔(图 2B-b、2B-c); 臀鳍较非生殖期增厚膨大, 但明显小于雌性(图 2B-b、2B-c), 解剖可见雄鱼腹腔内乳白色精巢。
2.2 东北七鳃鳗胚胎发育
东北七鳃鳗卵颜色为乳白色, 呈卵圆形(图3A), 长径和短径分别为(0.95±0.05)和(0.81±0.10)mm。卵受精后, 吸水膨胀, 受精膜举起(图 3B-a),受精膜外具有胶质膜, 有弱黏性。
卵裂期受精后6—8h, 在受精卵表面出现凹痕, 即分裂沟(图 3C-b), 受精卵开始第一次分裂。随分裂沟不断加深延长, 在中部环绕整个分裂球, 此时可见分裂球分成2个上下紧密相连、大小相似的细胞, 细胞呈半球形, 受精卵发育至2 细胞期(图 3C)。受精后8—10h, 分裂球进行第二次分裂,发育至4细胞期(图 3D), 此次分裂面与第一次分裂面呈直角, 在分裂球表面可见2条垂直的分裂沟, 分裂球均匀地分成4个等大的细胞。在4细胞期后约2h, 分裂球发育至8细胞期(图 3E), 在此期分裂球出现两种分裂方式: (1) 分裂球同时出现2条分裂沟(与第一次出现的分裂沟垂直), 分裂成8个细胞(排成2行, 每行4个); (2) 出现1条分裂沟, 分裂沟与第一次和第二次出现的分裂沟呈直角, 分裂球分裂成上下两层(每层各4个细胞)。随后, 胚胎约每2h进行一次卵裂, 依次进入16细胞期(图 3F)、32细胞期(图 3G)、64细胞期(图 3H)、128细胞期(图 3I)和256细胞期(图 3J)。
囊胚期受精后22—70h, 随细胞分裂次数的增加, 细胞明显减小, 细胞数量逐渐增加, 分裂沟不明显, 细胞间界线模糊, 整个胚胎发育成表面光滑的圆球型。在囊胚期初期, 胚胎内部形成较大的囊胚腔(胚胎下半部分的动物极呈半透明, 图 3K),随胚胎发育, 囊胚腔逐渐减小(动物极透明部分逐渐减少, 图 3L和 3M)。

图 2 繁殖期东北七鳃鳗的形态性状Fig. 2 Morphometric attributes of Lampetra morii at full sexual maturityA. 东北七鳃鳗雌鱼; a. 雌鱼(侧面); b. 背鳍、臀鳍和泄殖孔(侧面); c. 臀鳍和泄殖孔(腹面) B. 东北七鳃鳗雄鱼; a. 雄鱼(侧面);b. 背鳍、臀鳍和泄殖孔(侧面); c. 臀鳍和泄殖孔(腹面). 比例尺:1 cmA. L. morii female; a. side view of L. morii female; b. side view of dorsal fin, post-cloacal finfold and cloacal slit; c. ventral view of post-cloacal finfold and cloacal slit; B. L. morii male; a. side view of L. morii male; b. side view of dorsal fin, post-cloacal finfold and cloacal slit; c. ventral view of post-cloacal finfold and cloacal slit. Scale bars indicate 1 cm
原肠胚期受精后3—5d, 随胚胎发育, 细胞分裂次数增加, 数量继续增加, 个体不断减小, 原肠期胚胎植物极细胞被动物极细胞所包裹。胚胎植物极一端形成一拱形狭长的裂缝(图 3N-c)。随动物极细胞数量不断增加、移动和包裹, 胚胎体积减小, 胚孔不断内陷、扩大, 并逐渐向腹面迁移(图3O-c、3P-c和3Q-c)。
神经胚期受精后5—6d, 胚胎发育至神经胚期(图 3R、3S和3T)。此期胚胎胚孔背侧增厚延至胚胎前端, 背侧发育较平直, 胚孔迁移至胚胎的末端(图 3R-c和3T-c), 整个胚胎呈倒三角形。胚胎背部的外胚层不断增厚, 发育成神经基板, 神经基板中央下陷, 两侧外胚层隆起, 发育形成两条神经褶, 在胚胎背中线区域形成神经沟(图 3S-d和3T-d)。胚胎神经褶在背中线合并, 形成神经管。
头凸期受精后6—8d, 胚胎发育至头凸期(图 3U、3V和3W)。此期头部(图 3U-e和图3V-e)凸出于卵黄囊, 在头部腹侧可观察到凹陷, 咽部凸起(图 3U-f和图3V-f)。胚体增厚, 背部神经管(图 3U-g和图3W-g)明显凸起, 从头部延伸至胚体末端; 椭圆形的卵黄囊(图 3U-h、3V-h和3W-h)位于胚体后部, 胚孔(图 3V-c)迁移至卵黄囊末端。
孵出前期受精后8—11d, 胚胎发育至孵出前期(图 3X和图 3Y)。在早期(图 3X), 头部(图 3X-e)伸长, 并出现间歇性摆动。随胚胎发育, 头部和躯干部进一步拉伸, 胚体伸长, 胚胎在受精膜内卷曲(图 3Y)。在躯干部可见胚胎背侧肌节(图 3Y-i),卵黄囊由球型收缩成长条状(图 3Y-h), 头部间歇性摆动加快, 整个胚胎有规律地颤动和转动。
孵出期随胚胎生长和发育, 在受精后11—12d, 仔鱼孵出(图 3Z和图 3Z-1)。初孵仔鱼体色为乳白色, 身体未完全伸直, 后部弯曲, 呈现“J”型; 初孵仔鱼极少活动。在仔鱼头部(图 3Z-e和图3Z-1-e)腹侧可观察到鼻凹(图 3Z-j和图 3Z-1-j)和口凹(图 3Z-k和图 3Z-1-k), 两侧可见鳃原基(图 3Z-2-l)。在躯干部, 前端可见心脏(图 3Z-1-m)和肝脏(图3Z-1-n), 背侧可见神经管(图 3Z-1-g)和肌节(图 3Z-1-i)。卵黄囊(图 3Z-h和图 3Z-1-h)位于躯干部的中后部, 呈长条形棒状, 前细后粗。初孵仔鱼全长为(0.29±0.02) cm, 体重为(0.00032±0.00002) g。
2.3 卵黄囊期仔鱼生长和发育
卵黄囊期仔鱼的生长发育初孵仔鱼(0日龄)形态特征如2.2孵出期所描述(图 3Z、3Z-1和3Z-2)。1日龄仔鱼躯干后部及尾部向后拉伸, 身体较前一日平直, 仅靠近尾部分及尾部弯曲, 全长增加,整个身体透明度增加(图 4A)。在头部(图 4A-1-a)腹侧鼻凹(图 4A-1-b)和口凹(图 4A-1-c)加深、扩大, 鼻凹前移; 鳃原基增大(图 4A-1-d), 轮廓更加清晰, 鳃笼已具备雏形。在躯干部前端可见心脏膨大(图 4A-1-h), 有一心房一心室。躯干后部卵黄囊减小(图 4A-1-j), 卵黄开始吸收。身体背侧肌节明显(图 4A-1-g), 出现背鳍(图 4A-1-k)。在孵出后第2天, 仔鱼的全长增加, 尾部仍未平直, 鱼体透明度增加(图 4B)。头部背侧黑色素增加, 口和鼻凹陷加深并扩大, 鼻前移(图 4B-b), 鳃孔增大, 外部轮廓清晰。身体背侧肌节明显, 躯干部及尾部卵黄囊减小。3日龄仔鱼尾部向后伸展, 尾部与躯干基本平直, 仔鱼透明度增加(图 4C)。在头部背侧前端, 色素增加; 头部腹面口凹陷加深扩大, 鼻凹陷前移(图4C-b); 鳃孔轮廓清晰, 增大, 外部形态呈圆角方形。心脏开始博动, 在心脏内可见红色血液流动(图 4C-h)。随仔鱼的发育, 卵黄吸收, 卵黄囊减小。背褶发育, 出现尾鳍(图 4C-l)。在尾部腹侧可见肛门雏形(图 4C-m)。随仔鱼发育, 仔鱼(4日龄)的透明度增加(图 4D), 全长增长。头部口凹加深、扩大, 形成口笠(图 4D-1-n); 口笠内可见瓣膜(图 4D-1-r); 鼻凹陷向移至头部前端(图 4D-1-b); 眼点有少量色素沉着(图 4D-1-o); 鳃孔扩大, 轮廓清晰; 在第一鳃孔上方和眼的后方, 椭圆形耳囊形成(图 4D-1-p), 耳囊内可见耳石(图 4D-1-q)。在头部、躯干部体侧(卵黄囊的背侧)和背侧色素增加,呈“一”字型自前向后分布(图 4D-1-e)。背褶和尾鳍进一步发育。4日龄仔鱼开始上浮, 自由游动。如图 4E所示, 仔鱼(5日龄)全长增加。头部口笠进一步发育, 扩大, 口笠内可见质膜触手(图 4E-1-s); 鼻移至头部背侧; 眼点色素沉着, 形成一圆形黑点; 鳃笼发育, 鳃孔扩大, 轮廓清晰; 头部、躯干部体侧和背部色素明显增加。背褶和尾鳍膨大, 透明度增加。肛门形成一管状结构(图 4E-2-m)。6日龄仔鱼(图 4F)口笠扩大。鳃部发育, 主要表现为鳃笼增长,鳃孔增大。身体色素增加。肛门形成一管状结构与外界相通(图 4F-1-m), 其他形态较前一日基本相同。6日龄仔鱼钻入底层养殖沙中营穴居生活。随仔鱼的发育, 7—8日龄仔鱼透明度增加(图 4G、H)。口笠开口于头部前端, 口瓣开始活动。鳃孔增大, 鳃部轮廓清晰。仔鱼色素增加。随卵黄吸收,卵黄囊明显减小。其他形态特征与前一日基本相同。9日龄仔鱼(图 4I)的肝脏的前部出现胆囊(图4I-1-t), 其他形态特征与前一日基本相同。如图4J、4K、4L、4M和4N所示, 10—14日龄仔鱼色素增加。头部各组织和器官明显发育, 口笠扩大, 鳃笼生长明显, 主要表现为鳃孔增大, 轮廓清晰。10—14日龄仔鱼形态特征与9日龄基本相似。15日龄仔鱼(图 4O)的透明度增加, 体色加深。头部各组织和器官明显发育, 口笠扩大, 鳃笼生长明显, 主要表现为鳃孔增大, 轮廓清晰。卵黄囊完全吸收, 消化道贯通, 形成肠道, 开始滤食, 肠道充盈食物。
如图 5所示, 东北七鳃鳗卵黄囊期仔鱼的体重和全长均随日龄增加而增长。初孵仔鱼的体重和全长分别为(0.00032±0.00002) g和(0.29±0.02) cm,经15d卵黄囊期生长发育成幼鱼, 此时幼鱼体重和全长分别为(0.0012±0.0003) g和(0.80±0.06) cm。
卵黄囊期仔鱼身体各部分的异速生长东北七鳃鳗仔鱼在卵黄囊期发育过程中, 头长(图6A)、体长(图 6B)和尾长(图 6C)均表现出异速生长现象。仔鱼头长(1.421±0.102) mm和尾长(0.592±0.066) mm均在6日龄出现生长拐点。头长的异速生长指数在生长拐点前后分别为0.948 (P>0.05)和2.563 (P<0.05), 相对于全长生长速率分别表现为等速生长和正异速生长。尾长异速生长指数在生长拐点前后分别为3.373 (P<0.05)和2.176 (P<0.05), 尾长生长速率在生长拐点前后相对于全长生长速率均表现出正异速生长。而仔鱼体长在卵黄囊期内未出现生长拐点, 异速生长指数为0.352 (P<0.05),表现为负异速生长。

图 4 东北七鳃鳗卵黄囊期仔鱼外部形态特征Fig. 4 External view of Lampetra morii yolk-sac larva (head pointing left)A. 1日龄仔鱼; A-1. 1日龄仔鱼放大图; B. 2日龄仔鱼; C. 3日龄仔鱼; D. 4日龄仔鱼; D-1. 4日龄仔鱼放大图; E. 5日龄仔鱼; E-1. 5日龄仔鱼放大图; E-2. 5日龄仔鱼放大图; F. 6日龄仔鱼; F-1. 6日龄仔鱼放大图; G. 7日龄仔鱼; H. 8日龄仔鱼; I. 9日龄仔鱼; I-1. 9日龄仔鱼放大图; J. 10日龄仔鱼; K. 11日龄仔鱼; L. 12日龄仔鱼; M. 13日龄仔鱼; N. 14日龄仔鱼; O. 15日龄仔鱼. a. 头部; b. 鼻凹; c. 口凹; d. 鳃原基; e. 色素细胞; f. 神经管; g. 肌节; h. 心脏雏形; i. 肝脏雏形; j. 卵黄囊; k. 背鳍; l. 尾鳍; m. 肛门; n. 口笠; o. 眼; p. 耳囊; q. 耳石; r.瓣膜; s. 质膜触手; t. 胆囊; 比例尺: 1.0 mmA. 1-day old larva, A-1. enlargement of 1-day old larva, B. 2-day old larva, C. 3-day old larva, D. 4-day old larva, D-1. enlargement of 4-day old larva, E. 5-day old larva, E-1. enlargement of 5-day old larva, E-2. enlargement of 5-day old larva, F. 6-day old larva, F-1. enlargement of 6-day old larva, G. 7-day old larva, H. 8-day old larva, I. 9-day old larva, I-1. enlargement of 9-day old larva, J. 10-day old larva, K. 11-day old larva, L. 12-day old larva, M. 13-day old larva, N. 14-day old larva, O. 15-day old larva. a. head; b. nostril opening; c. mouth opening; d. gill anlagen; e. melanocyte; f. neurocoel; g. myomere; h. rudiment of heart; i. rudiment of liver; j. yolk sac; k. dorsal finflod; l.caudal finflod; m. rudiment of anus; n. oral hood; o. eye spot; p. otocyst; q. otolith; r. velum; s. oral cirri; t. gall bladder. Scale bars indicate 1.0 mm
卵黄囊期仔鱼头部器官的异速生长东北七鳃鳗卵黄囊期仔鱼的头部器官中, 吻长、眼径、眼鳃间距、口笠长、鳃前长和鳃长均随日龄的增加而增长, 但其生长速率相对于全长的生长速率并不相同。仔鱼吻长的生长拐点出现在9日龄[吻长:(0.595±0.055) mm, 全长: (6.532±0.305) mm], 生长拐点前异速生长指数为3.326 (P<0.05), 生长拐点后生长指数为2.271 (P<0.05), 均表现为正异速生长(图 7A)。仔鱼的眼径和眼鳃间距在卵黄囊期内并未出现生长拐点, 眼径异速生长指数为1.152 (P>0.05),表现为等速生长(图 7B); 眼鳃间距异速生长指数为0.687 (P<0.05), 表现为负异速生长(图 7C)。口笠长在12日龄出现生长拐点[口笠长: (0.315±0.023) mm,全长: (7.023±0.265) mm], 在生长拐点前其异速生长指数为0.870 (P<0.05), 表现为负异速生长, 而在生长拐点后其异速生长指数为2.888 (P<0.05), 表现为正异速生长(图 7D)。仔鱼的鳃前长在卵黄囊期内并未出现生长拐点, 其异速生长指数为1.546(P<0.05), 表现为正异速生长(图 7E)。仔鱼的鳃长在8日龄出现生长拐点[鳃长: (0.929±0.064) mm, 全长: (6.437±0.296) mm], 生长拐点前的异速生长指数为1.160 (P>0.05), 表现出等速生长; 而生长拐点后的异速生长指数为1.477 (P<0.05), 表现出正异速生长(图 7F)。
卵黄囊期仔鱼尾部器官的异速生长在卵黄囊期内, 东北七鳃鳗仔鱼泄殖孔长并未出现生长拐点。泄殖孔长的异速生长指数为0.589 (P<0.05),其生长速率相对于全长生长速率表现出负异速生长(图 8)。
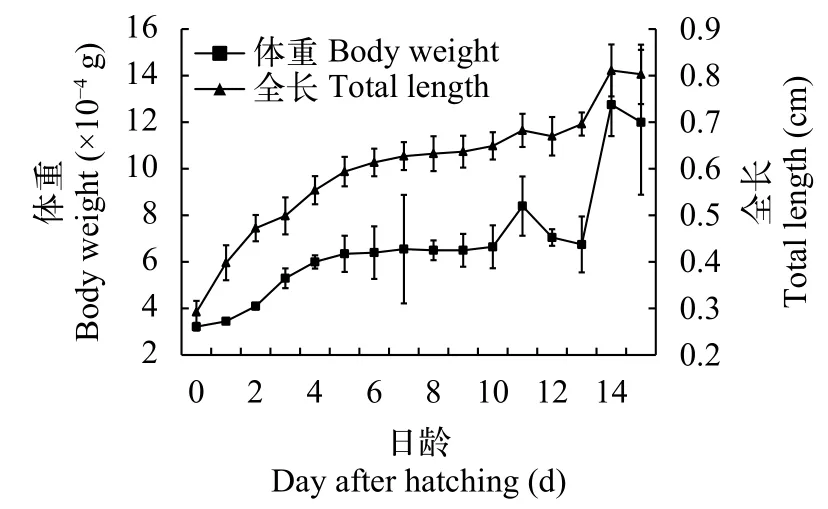
图 5 东北七鳃鳗卵黄囊期仔鱼体重和全长与日龄的关系Fig. 5 The daily body weight and total length of Lampetra morii yolk-sac larva
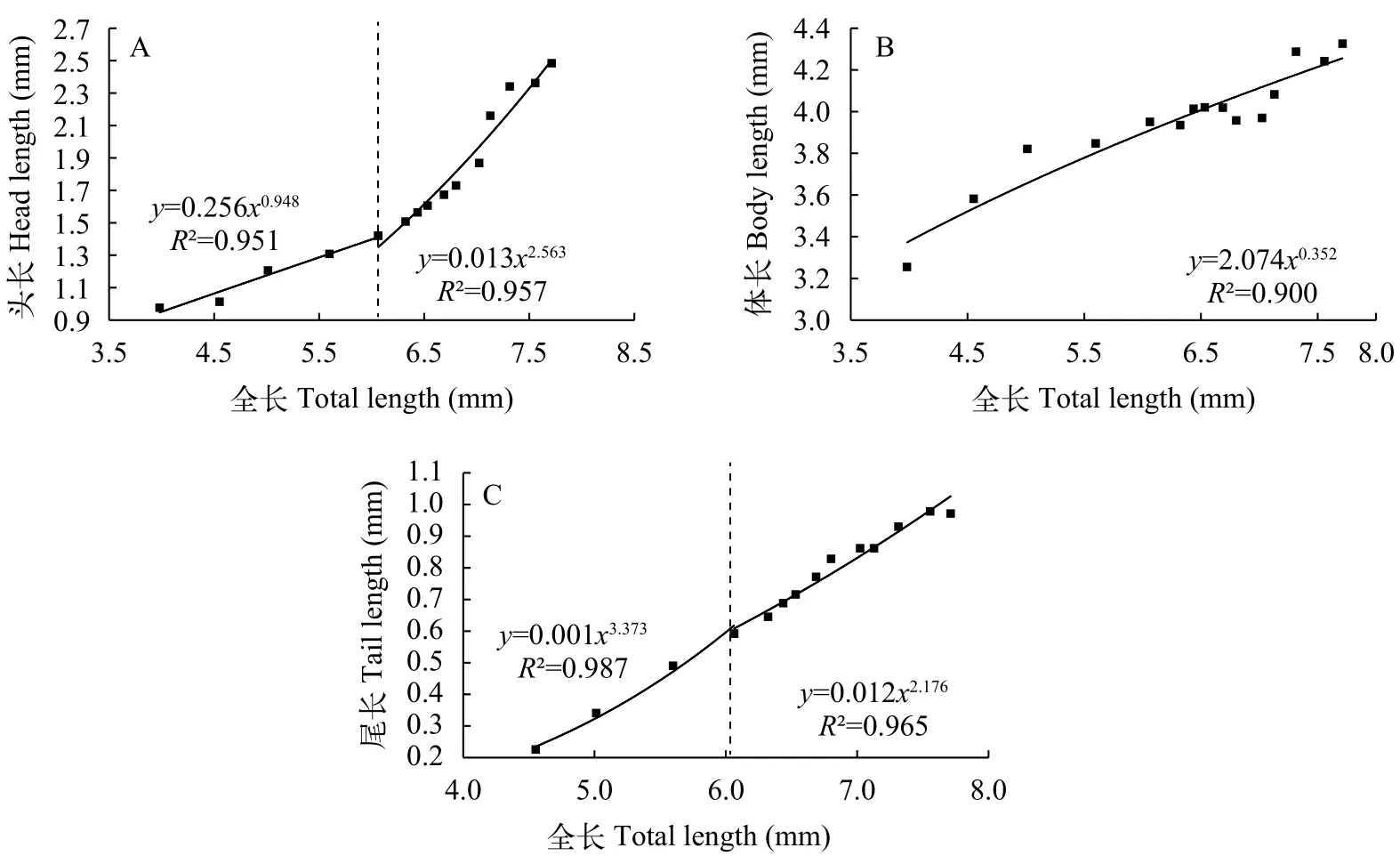
图 6 东北七鳃鳗卵黄囊期仔鱼身体各部分异速生长曲线及表达函数Fig. 6 Allometric growth curve and function of body parts of Lampetra morii yolk-sac larva
2.4 幼鱼生长发育
在1—5月龄幼鱼的发育过程中, 幼鱼大部分时间在缸底部的养殖沙中穴居, 活动能力较强, 其体色明显随月龄的增加而逐渐加深, 主要表现黑色素在幼鱼头部的口笠部、眼睛周围、鳃孔的周围和背侧, 躯干部体侧(侧线位置)和背侧、心脏的背侧以及尾部等部位的体表明显增多, 身体透明度逐渐减小。其他外部形态特征无明显的变化(图9 A—E)。
在幼鱼(1—5月龄)的生长发育过程中, 体重和全长均随日龄增加而增长。在15日龄时, 幼鱼的体重和全长分别为(0.0012±0.0003) g和(0.80±0.06) cm;经5个月的生长和发育, 150日龄幼鱼的体重和全长分别为(0.07±0.01) g和(3.87±0.32) cm (图 10)。
3 讨论
3.1 东北七鳃鳗副性征和繁殖行为
副性征是指性成熟动物外表所显示出的雌雄特征[17], 具有季节性变化, 是性腺发育的外观特征,成为其鉴别雌雄的主要依据。本研究表明, 在繁殖季节室内暂养的东北七鳃鳗雌性和雄性个体显示出副性征, 与海七鳃鳗[6,9,18]、雷氏七鳃鳗(Lampetra reissneri)[19]、Lampetra lamptteni[17]、Ichthyomyzon fossor[17]、Ichthyomyzon castaneus[17]和Ichthyomyzon unicuspis[17]相似。但不同七鳃鳗物种的副性征并不相同, 具有各自特点, 如性成熟海七鳃鳗在鳃孔后方的背部至第一背鳍出现一“线状组织”[16]; 性成熟I. castaneus[17]和I. unicuspis[17]的泄殖孔并未出现肿胀和充血现象。
以上研究表明, 东北七鳃鳗成体在室内暂养越冬后, 在繁殖季节显示出副性征和繁殖行为, 表明其在室内越冬的可行性, 为东北七鳃鳗的人工养殖奠定了基础。同时, 为七鳃鳗的繁殖生物学研究和人工繁殖提供了参考依据。

图 7 东北七鳃鳗卵黄囊期仔鱼头部各器官的异速生长曲线及表达函数Fig. 7 Allometric growth curve and function of head organs of Lampetra morii yolk-sac larva
3.2 东北七鳃鳗的早期发育
卵径是反映动物生活史进化过程的一个重要的生物学特征, 其大小与环境相关, 是长期适应环境的结果。已有研究报道溪水型七鳃鳗卵径大于洄游型七鳃鳗卵径[20,21]。本研究表明, 东北七鳃鳗卵径[长径为(0.95±0.05) mm、短径为(0.81±0.10)mm]大于同一地区洄游型日本七鳃鳗卵径(长径和短径分别为0.846和0.759 mm[8])。Yamazaki等[21]认为, 七鳃鳗表型特征应向小体型(繁殖期)、大卵径和低繁殖力的生物特征进化, 这可能与七鳃鳗的产卵场环境恶劣有关(如饵料缺乏、水温低、水流速快等), 体积相对较大的卵能获得更大的生存机率。
本研究参考海七鳃鳗[6]、E. tridentatus[7]、日本七鳃鳗[8]和L. reissneri[19]外部形态特征划分东北七鳃鳗胚胎发育时期。研究表明, 东北七鳃鳗卵形态、卵裂方式、胚胎发育和卵黄囊期仔鱼发育与海七鳃鳗[6]、E. tridentatus[7]、日本七鳃鳗[8]和L.reissneri[19]相似。曲漱惠等[22]认为划分胚胎发育时期不仅依据其外部形态特征, 还应结合胚胎内部组织结构变化特点。李军等[8]研究日本七鳃鳗胚胎发育过程中的外部形态和内部组织结构, 确定胚胎在64细胞期和128细胞期分别进入囊胚期和原肠期。这与本研究结果并不相同, 所以在下一步研究工作中, 结合东北七鳃鳗各胚胎发育时期内部组织结构,开展东北七鳃鳗胚胎发育研究。
东北七鳃鳗初孵仔鱼全长(0.29±0.02) cm与日本七鳃鳗(0.30±0.03) cm[23]和海七鳃鳗[6](0.3—0.5 cm)相近; 而其体重(0.00032±0.00002) g大于日本七鳃鳗(0.00016±0.00003) g[23]。经过卵黄囊期, 东北七鳃鳗幼鱼较日本七鳃鳗[23]的个体大, 卵黄囊期仔鱼生长速率(0.034 cm/d、0.000059 g/d)高于日本七鳃鳗(0.027 cm/d、0.000035 g/d)。在幼鱼期, 经5个月的生长和发育, 与在同一条件下室内养殖的日本七鳃鳗[23]相比, 东北七鳃鳗幼鱼同样表现出生长速率快, 个体较大。与野外采捕的1龄L. planeri和海七鳃鳗个体(全长3—4 cm)[16]相近。东北七鳃鳗早期发育阶段生长速率与物种、栖息地理微环境以及幼鱼的密度等密切相关[16], 同时也可能与室内良好的养殖水质, 相对较高的水温以及丰富、营养均衡的饵料有关。东北七鳃鳗的早期发育研究不但为七鳃鳗的发育学研究提供了基础数据, 还显示东北七鳃鳗是适合室内养殖的优良物种。

图 8 东北七鳃鳗卵黄囊期仔鱼泄殖孔异速生长曲线及表达函数Fig. 8 Allometric growth curve and function of cloacal length of Lampetra morii yolk-sac larva

图 9 东北七鳃鳗幼鱼外部形态特征Fig. 9 External view of Lampetra morii ammocoeteA. 30日龄幼鱼; B. 60日龄幼鱼; C. 90日龄幼鱼; D. 120日龄幼鱼; E. 150日龄幼鱼. 比例尺: A—B. 1.0 mm; C—E. 1.0 cmA. 30-day old ammocoete, B. 60-day old ammocoete, C. 90-day old ammocoete, D. 120-day old ammocoete, E. 150-day old ammocoete.A—B scale bars indicate 1.0 mm; C—E scale bars indicate 1.0 cm
3.3 东北七鳃鳗卵黄囊期仔鱼的异速生长
在水生生物的早期发育过程中, 组织形成和器官分化是极其重要而又复杂的过程, 其在外部形态、新陈代谢以及生活行为上的变化是生物体为适应外界环境做出的选择[24]。在七鳃鳗早期发育过程中, 仔鱼的感觉、摄食、呼吸和游泳器官的异速生长是长期进化过程中保留下来的发育特征[8],目的是使仔鱼能够在最短的时间内获得早期生存的各项能力。
卵黄囊期, 东北七鳃鳗仔鱼头长和尾长的生长速率相对于全长均表现为正异速生长, 而体长则为负异速生长, 与日本七鳃鳗仔鱼的异速生长现象相似[8]。七鳃鳗头部的正异速生长, 可以更好地保证组织和器官发育, 如大脑、视觉、呼吸和摄食器官。其次, 头长和尾长的正异速生长能增强仔鱼的游泳能力, 而体长的负异速生长则能有效地控制七鳃鳗的全长, 减小头部与尾部间的距离, 有利于游动中身体平衡[25]。东北七鳃鳗头长和尾长出现生长拐点时间(6日龄)与日本七鳃鳗出现生长拐点的时间相近[8], 这可能与其开始穴居生活方式有关[8]。
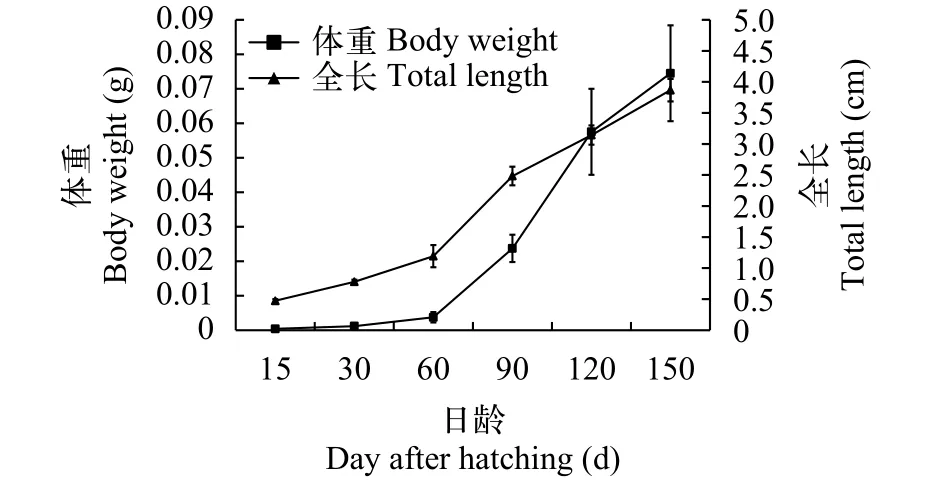
图 10 东北七鳃鳗幼鱼体重、全长与日龄的关系Fig. 10 The body weight and total length of Lampetra morii ammocoete
口笠是仔鱼的摄食器官, 直接决定了滤食能力的强弱。东北七鳃鳗的口笠的异速生长与日本七鳃鳗相似[8], 在生长拐点后呈正异速生长。口笠的发育与吻长和鳃前长关系密切, 吻长和鳃前长快速发育为口笠的生长提供了适当的空间[8]。而东北七鳃鳗在卵黄囊期泄殖孔表现的负异速生长现象则与日本七鳃鳗不同。
东北七鳃鳗鳃长的异速生长与日本七鳃鳗相同[8]。东北七鳃鳗初孵仔鱼在水底生存, 约在6日龄开始穴居生活。鳃长在8日龄出现生长拐点, 生长拐点后鳃长的正异速生长速度高于生长拐点前。鳃的正异速生长加快了鳃的发育, 能够保证仔鱼获得足够的溶解氧, 这可能与其穴居生活方式有关[8]。眼鳃间距在卵黄囊期一直处于负异速生长, 为鳃的正异速生长提供了足够的空间[8]。
七鳃鳗仔鱼和幼鱼主要在水底和底层泥沙中营穴居生活, 其栖息环境较暗, 以被动滤食腐植碎片、有机碎屑和浮游生物等为生, 眼睛的发育对于七鳃鳗仔鱼的早期生存作用较小, 这可能是东北七鳃鳗和日本七鳃鳗眼径呈负异速生长的主要原因[8]。
综上所述, 东北七鳃鳗卵黄囊期仔鱼发育过程中存在异速生长现象, 是其适应环境的重要生长策略。
