杉木活立木组织内的养分转移特征
2019-07-09周玉泉康文星陈日升田大伦项文化
周玉泉,康文星,2,3,陈日升,田大伦,2,3,项文化,2,3
(1.中南林业科技大学,湖南 长沙 410004;2.南方林业生态应用技术国家工程实验室,湖南 长沙 410004;3.湖南会同杉木林生态系统国家野外科学观测研究站,湖南 会同 418307)
植物组织内的养分转移属于养分内循环。这种循环有两种过程,一是植物活组织之间的养分转移[1],二是植物组织衰老时养分向活组织转移[2-4]。以往植物养分内循环研究,几乎全部集中在衰老组织养分向活组织转移上。如不同生活型植物衰老叶片养分转移率的差异[5-8],不同林龄植物衰老叶片养分转移的变化规律[9-11],不同气候和气候变化对植物叶衰老时养分转移的影响[12-14],衰老叶片养分转移率与土壤肥力的关系,施肥以及温室条件下养分有效性对养分转移的影等响[15-16]。然而,到目前为止,国内外却鲜见植物活组织之间的养分转移的研究报道。早在1977年,有学者就提出养分元素在树体内的活动性思想[17],Miller[18]根据这种思想对养分元素在树体内的运转和位移作了总结。实际上植物的正常生长发育过程中,植物活组织内养分流动普遍存在[19]。如何估算活立木组织内的养分转移量,一个林分从幼林到成熟期不同年龄阶段活立木组织内的养分转移规律怎样?这些是森林养分循环深入研究中亟需解决的问题。本研究利用会同杉木林25 a 定位测定的生物量和养分数据,根据某林龄段前后两次测定的现存生物量和枝、周期估算某林龄段林分生产的生物量;用某林龄段前后两次测定的养分浓度数据,估算某林龄段的以前生长的,在这林龄段还存活的生物质中新补充或转移出的养分量,探讨不同林龄杉木活立木组织内养分转移的过程和规律。为人工林养分循环的研究和经营管理提供科学依据。
1 研究区概况
本研究在湖南会同杉木林生态系统国家野外科学观测研究站第3 试验小集水区的杉木人工林内进行。地理位置 26°50′N ,109°45′E 。面积1.984 hm2,海拔270~350 m,坡向西北,坡度15 °~25 °。研究的林分是1977年冬采伐第一代杉木人工林后,1988年春在采伐迹地上采用炼山、全垦挖穴营造的第2 代杉木林。造林密度约2 200 株·hm-2。杉木栽上后前3年的春秋季节各抚育一次,以后任其自然生长。
研究区属于中亚热带季风湿润气候区,年平均气温16.9 ℃,年降水量1 100~1 400 mm,年均相对湿度80%以上。土壤母岩为震旦纪板溪系灰绿色板岩,风化程度较深,土层厚度80 cm 以上,土壤为黄壤。A1 层腐殖质含量2%~4%,C/N 值12~14,pH 值4.8~5.7。林地45 cm 深层土层平均养分浓度:有机质3.027%~4.859%,N 1.36~2.03,P 0.48~0.65,K 13.65~21.27 g·kg-1。林下植被主要有杜茎山Maesa japonica、山苍子Litsea cuheha、白桤木Pinus albicaulis、白背叶Mallotus apelta、 冬 青Ilex chinensis、鱼腥草Houttuynia cordata、狗脊蕨Woodwardia japornica、仙鹤草Herba agrimoniae、梵天花Urena procumbens等组成。
2 研究方法
2.1 样品采集
第2 代杉木人工林7年生时,设立了4 块固定样地(每个样地面积为667 m2)并根据原林业部科技司主编的《森林生态系统定位研究方法》[20]提供的克拉夫特的林木生长状况分级法测定林分生物量。以后在林分11、16、20 和25年生时,都在4 块样地测定林分生物量。每次测定生物量的同时采集养分待测样品。为了保证样品具有代表性,将每一次采集样品伐倒的标准木(依据样地的平均树高、平均胸径测定的标准木)分成树干基部、中下、中、中上和上部5 个区段,每一区段釆集相同质量的树干,再把釆集的树干混合一起组合成树干样品。 树皮取样采用树干相同的方法。枝和叶都分成当年生、1年生、2年生、多年生分别取样。根分成根头、大根(1 cm ≤根径)、粗根(0.2 cm ≤根径<1 cm)、细根(根径< 0.2 cm)分别取样(杉木各器官取样样本分别为2 kg 左右)。
2.2 养分测定
将待测的杉木各器管组织样本,取适量样品置于85℃恒温干燥箱内烘干至恒质量。称干质量,磨碎,过60 目筛后按《土壤农业化学分析方法》[21]配置待测溶液。用半微量凯氏法测定植物中的全N,用分光光度计测定植物中全P,用原子吸收分光光度计测定植物中全K、全Ca、全Mg。每个样品重复测定3 次,用3 次重复测定的平均值作为计量标准。
2.3 不同林龄林分养分积累现存量计算
林分某一林龄时养分积累现存量(M)用下式计算:

式(1)中:Wi为林分某一林龄时杉木某器官现存生物量(t·hm-2);Li为林分某一林龄时杉木某器官的养分含量(g·kg-1);i=1,2,…,5(干、皮、枝、叶、根)。
2.4 某林龄阶段开始时的生物现存量在某林龄段内仍存活的生物量估算
根据杉木干、根、皮在生长过程中不发生枯死现象(因很难准确测定根的生长周期,大多数研究者都将根与干材和树皮同等对待),因此某个林龄段开始时的干、根、皮现存生物量在这个林龄段内不会发生变化。杉木枝叶有一定的生长周期(本研究中杉木叶的生长期定为5 a[22],根据在会同对杉木生长的长期观测,把枝生长期定为10 a),枝叶在生长过程中不断代谢凋落,也就是说某个林龄段开始时的枝、叶现存生物量,有些会在这个林龄段内枯死凋落而发生变化。本研究利用陈日升等[23]的研究方法,估算某个林龄段开始时的林分现存生物量在这个林龄段内还活着的量。
2.5 某林龄段以前生产的且在某林龄段还存活生物量中的养分转移量计算
根据某林龄开时的杉木各器官养分含量(L1m)和结束时的含量(L2m),以及某林龄段以前生产的在某林龄段还存活生物量W6m,估算某林龄段内这些还存活的生物量的养分转移量。若L1m>L2m时,意味某林龄段以前生长的且在这林龄段还存活植物组织养分含量降低了,有部分养分被转移出来。转移量M2计算公式为:

当L1m<L2m时,表示某林龄段以前生长的且在这林龄段还存活植物组织养分含量升高了,有新的养分补充转移到这些存活各器官内。补充转移量M3计算公式则为:
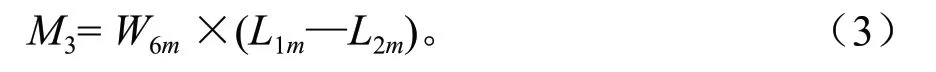
2.6 植物体内养分转移率计算
根据养分转移率定义(转移的养分量与能够进行转移养分总量的比值),如果某林龄段以前生长的且在这林龄段还存活植物组织内转移出的养分,其养分转移率(F)为:

式(4)中:M2见公式(2);M4为某林龄段以前生长的,且在这林龄段还存活植物组织内的养分总量,其计方法:

式(5)中:Li表示某林龄段以前生长的且在这林龄段还存活植物组织养分含量(g·kg-1);i=1,2,…,5(干、皮、枝、叶、根)。
2.7 数据分析处理
所有数据经 Excel 2003 软件处理,用SPSS 20.0 软件进行统计分析。采用单因素方差分析(one-way ANOVA) 比较不同生育阶段,不同器官养分流动量的差异显著性,显著水平为P= 0.05。
3 结果与分析
3.1 某林龄阶段开始时的生物现存量在某林龄段内仍存活的生物量
7年生时测定的林分生物量,实际是从林分造林那年到第7年生产的生物量。这个林龄阶段以前生长的,在这林龄段还存活植物生物量实际上不存在。所以表1中只有8年生后各林龄段以前生长的,在这林龄段还存活生物量数据。从表1中看出,某林龄段开始时的林分生物现存量,在这林龄段内随着林龄段内的林龄增加逐渐减少。在8~11,12~16,17~20 和21~25 a各林龄段,某林龄段开始时的林分生物现存量,那些在这林龄段内仍存活的生物现存量年均分别为36.01,66.76,104.94,147.16 t/hm2。 与 某 林龄段开始时的现存生物量相比分别减少9.77%~11.75%。
从表1中还看出,某林龄段开始时的树干、皮和根生物现存量在某林龄段内其现存量没有变化,主要是枝和叶的生物现存量减少了。这是因为杉木干、根、皮在生长过程中不发生枯死现象,而枝叶具有一定的生长周期引起的。例如12~16 a 林龄段,这林龄段的年份数为5年恰好为叶的生长周期,那么林分12年生以前生长的叶在这林龄段结朿那年都会枯死,所以到林分16年生时,林分12年生以前生长的叶现存量为零。
3.2 某林龄段以前生产的且在某林龄段还存活生物质中的养分转移量
同样由于1~7 a 这个林龄阶段以前生长的,在这林龄段还存活植物生物量实际上不存在。所以表2和表3中只有8 a 后各林龄段的以前生长的,在这林龄段还存活植物组织的养分转移数据。

表1 某林龄段的以前的生物现存量在这林龄段内仍存活的量†Table1 Amount of previous living biomass of a forest age segment that survives in this age group (t·hm-2)
从表2看出,8~11 a,转移到以前生长的,且在这林龄段还存活植物组织内的新的N、P、K、Ca、Mg 量,年均分别为3.11,0.37,1.36,1.96 和1.03 kg·hm-2a-1。11年生后各林龄段,某林龄段以前生长的,在某林龄段还存活植物组织内转移出的N、P、K、Ca、Mg 量,年均分别为1.30~1.66,0.40~0.53,1.06~1.40,1.77~3.26,0.65~0.90 kg·hm-2a-1。林分11年生后,养分元素转移量随着林龄增加变化为单波峰曲线。因为11~20 a 时,林分处于干材生长和近熟林阶段,杉木各器官功能旺盛,因而从以前生长的在这林龄段还存活植物组织内,运转出的养分量逐年增加;进入成熟期后(20~25 a),杉木各器官功能减弱,导致以前生长的在这林龄段还存活植物组织内运转出的养分量逐年减少。
差异检验表明,相同林龄段各养分元素运转量差异显著(P<0.05)。不同林龄段的相同元素运转量除12~16 和20~25 a 的P 与P之间,8~11 和12~16 a 的Ca 与Ca 之间差异不显著外(P>0.05),其差异显著(P<0.05)(表2)。

表2 不同林龄阶段杉木林树木组织内的年均养分元素转移量†Table2 Average annual transfer of nutrient elements in the tissues of Chinese fir forests at different age stages (kg·hm-2a-1)
由表3可知,8~11 a年平均输送到原来生长的,在这林龄段还存活的树干、皮、枝、叶和根的新的养分量,分别为1.01,1.21,2.10,2.92和0.59 kg·hm-2a-1。可见,在幼树到快速生长阶段,生理功能越大的器官补充的养分越多。11年生后各林龄阶段,从原来生长的在某林龄段还存活的树干、皮、枝、叶和根中,年均转移出的养分量依次是1.09~1.79,0.64~0.87,1.08~1.63,0.86~1.57,0.88~1.28 kg·hm-2a-1。各器官转移送出的养分随林龄变化规律是:除从叶中转移出来养分量的随林龄增加而减少外,树干、皮、枝和根中转移出来的养分量的随林龄增加的变化呈现先上升后下降的趋势。而且当杉木进入近熟期(17年生)后,从树干和树枝中转移出来的养分量大于其它器官。可见,各器官输送出来的养分量也与器官的功能有关,进入近熟期后,制造有机物质和运送养分的器官如树皮、树叶转移出来的养分较少,那些生理功能不大的器官如从树干、枝中转移出来的养分较多。

表3 不同林龄段杉木林树木器官的年均养分转移量Table3 Average annual nutrient transfer of tree organs in Chinese fir forests of different age groups (kg·hm-2a-1)
差异性检验表明,除12~16 a 树干与树根,17~20 a 树干和树枝、树叶与树根之间,21~25 a 树叶与树根之间转移的养分量差异不显著外(P>0.05),其余同龄的各器官转移的养分量差异显著(P<0.05);不同林龄同一器官转移的养分量除17~20 和21~25 a 的树干之间差异不显著外(P>0.05),其余差异显著(P<0.05)(表3)。
3.3 某林龄段以前生产的且在某林龄段还存活生物质中的养分转移率
表4看出,11年生后各林龄段,由某林龄段的以前生产的,且在某林龄段还存活生物质中转移出的各养分元素转移率的大小顺序是: P(3.69%~6.04%)>Mg(2.59%~5.02%)>Ca(2.02%~3.38%)>K(1.41%~2.91%)>N(1.10%~2.09%)。可见,各养分元素的转移率被此不相等。各养分元素的转移率的大小可能与养分元素在植物体内的功能作用有关。P 磷元素参与卵磷脂、细胞核蛋白的构成,能加速细胞分裂,促进花芽分化,促使根系和地上部加快生长;Mg 是多种酶的活化剂,可促进植物生长发育,且植物体内70%的Mg 以游离态存在属于较易移动的元素。所以这两种元素的转移率较其它元素大。
从表4中还看出,各养分元素转移率及总的养分转移率随林龄增加而降低。因为养分转移率定义为转移的养分量与能够进行转移养分量的比值,也就是某林龄段以前生长的,且在这林龄段还存活植物组织内转移出的养分与某林龄段以前生长的,且在这林龄段还存活植物组织内养分储量的比值。虽然,某林龄段以前生长的,且在这林龄段还存活植物组织内转移出的养分随林龄变化为抛物线,但某林龄段以前生长的,且在这林龄段还存活植物量随林龄增大而增加,因而其养分储量随林龄增大而增大。而且其养分储量随林龄增大的速率高于同林龄期从这些物质中转移养分量的增加速率,因此,导致各养分元素转移率及总的养分转移率随林龄增大而降低。
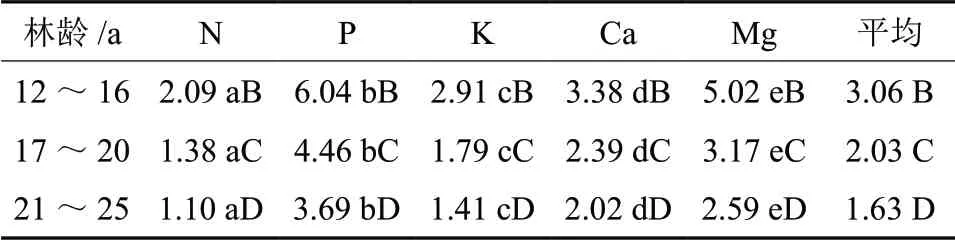
表4 某林龄段以前生产的且在某林龄段内仍存活生物质中转移出的养分的转移率Table4 Transferred rate of nutrients from biomass produced previously in a forest age segment and is still alive in a forest age segment %
差异检验表明,相同的林龄,各营养元素转移率差异显著(P<0.05);不同林龄的同一营养元素转移率差异也显著(P<0.05)。
4 结论与讨论
4.1 与植物组织衰老时养分转移的差异
目前还未见有关活立木组织之间的养分转移的研究报道,但植物组织衰老时向活组织转移养分的报道颇多。因此,本研究只能与植物组织衰老时向活组织转移养分的有关研究进行对比分析。衰老植物组织的养分转移方向是由衰老组织到活组织。活立木组织内养分转移方向受养分含量差异制约。若某林龄段结朿时杉木体内养分含量大于某林龄段开始时,养分转移方向为新的养分输入到在某林龄段的以前生产的,且在某林龄段还存活物质中;当某林龄段结朿时杉木体内养分含量少于某林龄段开始时,则是某林龄段以前生产的,且在某林龄段还存活物质中的养分被转移出来。而且,活立木组织内养分转移方向还受林木不同生育阶段特征的影响。杉木幼林和快速增长阶段(林分11年生以前),由于要壮大自己的个体,必需从土壤中吸收大量养分充实到某林龄段的以前生长的,且在某林龄段还存活的物质内,其养分流向由从土壤吸收的养分流向以前生长的,且在某林龄段还存活植物组织内体。当林分郁闭后,生产单位生物量所需养分量下降,体内养分再分配机制导致积累在某林龄段的以前生长的,在某林龄段还存活的物质内的养分转移出来用于新的物质生产[24]。本研究中11年生以后各林龄段(林分已郁闭),其养分流向由某林龄段的以前生产的,且在某林龄段还存活物质中的养分被转移出来再利用。
许多研究表明,衰老叶片中有部分N、P、K向活体发生转移,Ca、Mg 的移动性差或为不发生转移[5-8,25],这与本研究结果稍有不同。本研究中N、P、K、Ca、Mg 都发生转移。衰老叶中的N、P、K 的转移率都较大[8,25-27],本研究中由某林龄段以前生产的,且在某林龄段还存活的植物组织内转移出的各营养元素转移率最大不超过7%。本研究中立木活组织之间养分的转移方向、转移量和转移率与植物组织衰老时的养分转移到活组织有较大差别,这是两种内循环的生理过程和调控机制不相同引起的。但本研究中当林分郁闭后,某林龄段以前生产的,且在某林龄段还存活植物组织内,转移出的营养元素转移率随林龄增加而减少,这与曾德慧等[28]、邓浩俊等[11]研究的叶在枯死时养分元素转移率随林龄增加而下降的结论相同。表明植物两种内循环功能的发挥都与林木生长发育规律有关。
4.2 杉木活立木组织内养分的转移量及意义
活立木组织内养分转移量大小也与养分含量差异和器官生理功能有关。显然,养分含量差异越大,驱使养分流动的作用力也越大,转移的养分也就多。如本研究11年生与7年生杉木体内养分含量差异大于16年生与11年生的[23],所以8~11 a 的养分转移量也就比12~16 a 多。叶制造有机物质是功能最活跃的器官,所需养分量最多,树干主要起着植物体的支撑作用,其生理活动较弱,所需的养分较少,因此,重新补充到原来生长还依然存活的植物各器官的新的养分,叶中最多,干中最少。同样,从某林龄段以前生长的,且在某林龄段还依然存活器官,转移出来的养分量也与器官生理功能相关。那些生理功能作用大,生命活动激烈的器官如树叶,需要较多的营养物质才能维持他们的功能正常发挥,因此,从这些器官转移出来再重新利用的养量不会很多。干、枝起着植物体的支撑作用,其功能作用没有其它器官那么重要,所以才能有较多的养分转移出用来再利用。
活立木体内的养分转移加强了森林养分的循环[29];在维持植物体内化学元素计量平衡中也起着重要作用[31];这些养分可以直接用于植物生长,提高了养分的利用效率,减少了植物对土壤养分的吸收,这对林地地力的维护有重要意义[1,31]。林木体内转移的养分的再利用是植物养分利用策略的重要驱动力,降低对环境的依赖性。所以,林木体内转移养分再利用的变化对生态系统养分循环、植物种群和群落稳定性都具有重要的意义。
4.3 活立木体内养分转移的机理探讨
有研究表明,从土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中植物[32],土壤贫瘠的胁迫作用可能是活立木体内养分转移的机理。也有研究指出,植物养分转移与土壤养分供给性大小相关性不大,贫瘠土壤并不一定是植物体内养分转移的驱动力[26,33-35]。目前比较一致的认为是植物体内养分的转移和再利用只是植物对养分贫瘠环境的一种适应机制,也是植物保存养分并维持体内养分平衡的一种重要养分利用策略[36-39]。一些研究者用植物生理中“源-汇”理论分析树木调节体内的养分转移的生理机制。认为树木器官在某些生理条件下是养分的源,在另一些生理条件下可能是汇。正由于这种汇源的作用,调控树木体内的养分流动[40-41]。如本研究中,杉木幼树和快速增长阶段由于要壮大自己的个体,必需从土壤中吸收大量养分充实到某林龄段以前生长的,且在某林龄段还存活各器官组织内。各器官组织成了养分的汇。当林分郁闭后,积累在某林龄段以前生长的,且在某林龄段还存活的物质内的养分转移出来再利用,各器官组织成了养分的源。有学者认为活立木体内养分转移的机理是植物本身具有这种自我调控的反馈机制,当植物体不同部位或组织的养分达不到平衡时,通过这种调控机制使养分在活体内转移来维持体内养分平衡,而这种调控机制是高大乔木在自然界长期的生存竞争中形成的[24]。树木体内的养分循环是一个非常复杂的生理生化过程,目前对活立木体内养分转移控制机制还缺乏基本的了解,对它们的形成机制的研究仍是一个非常薄弱的环节,揭示活立木体内的养分转移内在调控机制仍然是生理生态学亟待解决的重要问题。
4.4 研究中存在的问题及今后研究的方向
本研究由于没有估算不同林龄段杉木林的土壤养分吸收量,因而没有得出重新补充到某林龄段以前生长的,且在某林龄段还存活的物质内的新的养分的转移率(因为新补充的养分来自土壤养分)。而且本研究只对某林龄段以前生长的,且在某林龄段还存活的物质内的养分转移动态进行分析,并没涉及某林龄段新生产物质内的养分流动过程,因而还无探明植物养分体内养分迁移的内在机理。
对植物体内养分内循环的研究,可以追溯到20 世纪30年代,科学家们就已经注意到叶片养分在衰老过程中向活组织转移的现象[42]。到目前仍有大量养分在植物组织想衰老过程中向活组织转移的研究报道[43-45]。但对于植物体内活组织间的养分迁移,直到20 世纪50年代初,Gregory 利用同位素示踪技术首次在禾谷类草本植物中发现了养分在植物体内活组织间的迁移现象[46]。由于实验条件(方法)的限制,目前同位素示踪法只用于草本植物体内养分内循环研究[45]。迄今为止,国内外还未见利用同位素示踪法用于多年生乔木的养分内循环研究报道。今后应采用先进的技术和手段研究植物体内养分内循环,只有这样才能更好地开展植物养分的生物化学循环研究,才能更好地探明植物养分内循环的内在机理。而且今后应把树木个体水平的信息应用同位素技术转移到生态系统水平,应利用同位素示踪技术把植物体内养分内迁移效率和养分利用效率结合一起研究,这样才能为植物养分利用效率和生产力的提高提供科学依据。
