基于同步辐射显微CT研究冻融循环对黑土团聚体结构特征的影响
2019-07-08范昊明侯云晴郭芯宇马仁明
姜 宇,范昊明,侯云晴,刘 博,郭芯宇,马仁明
沈阳农业大学水利学院,沈阳 110866
冻融作用作为一种自然现象普遍存在于中、高纬度及高海拔地区。东北黑土区位于中纬度地带,秋末和春初易形成土壤季节性融化层与冻土层的周期性变化[1-2],该周期性变化对土壤结构产生强烈的影响。土壤结构指土壤中原生颗粒和次生颗粒以及土粒间所构成孔隙的不同排列形式,是维持土壤功能的基础,也是影响侵蚀过程的重要因素。作为土壤结构重要组成单元的团聚体,其大小分布和稳定性影响着土壤的孔隙性、持水性、通透性和抗蚀性[3-4]。因此,研究冻融作用对黑土团聚体结构特征的影响具有重要意义。
国内外关于冻融循环对团聚体影响的研究已经逐步开展,由于冻融作用对土壤结构会产生影响,因此,冻融过程被认为是影响团聚体形成和破碎的重要因子。Benoit研究发现冻融作用对团聚体破碎的影响与冻融强度、团聚体颗粒大小和团聚体水分含量有关[5]。Six则表示冻融过程对团聚体的物理破坏随冻融次数不断累积,其中大团聚体破碎尤为明显[6]。此外,关于冻融作用对团聚体影响的研究多侧重于其对团聚体稳定性的影响。王恩姮等[7]、范昊明等[8]在此方面进行研究得出相近的结论,即冻融作用使大团聚体的稳定性降低,但提高了小粒级团聚体的稳定性。
前人对于冻融作用下团聚体的形成、破碎机制及稳定性变化做了大量研究,但缺乏对团聚体内部孔隙特征的研究,而团聚体孔隙特征决定了团聚体的结构性,因此,开展冻融作用对团聚体孔隙特征影响的研究尤为重要。随着技术的发展,高精度同步辐射CT技术对土壤孔隙结构的可量化研究已达到微团聚体尺度[9-11]。我国上海光源X射线成像及生物医学应用光束线站(BL13W1)可以进行高分辨率的三维成像,为三维土壤微结构研究提供了条件。本研究利用上海光源同步辐射显微CT对冻融作用下5—7 mm的大团聚体的三维结构进行分析,获取孔隙特征的定性、定量指标,进而分析不同冻融循环周期对土壤大团聚体孔隙结构特征的影响。为进一步揭示黑土区季节性冻融对黑土结构的影响以及水土流失的防治提供科学理论依据。
1 材料与方法
1.1 试验点与样品采集
取土地点为黑龙江省齐齐哈尔市拜泉县,地理坐标为126°18′43.7796″E,47°27′42.0726″N。用内径为4.8 cm,高为15 cm的PVC管采集0—15 cm土层范围的原状土体。采样方式为原位静压法,采集后将土柱的上下两端均用保鲜膜封闭,防止土壤水分快速散失发生干裂;在采集、运输和试验过程中注意防止对原状土体结构的扰动。
冻融试验前,将原状土柱置于4℃下恒温保存。利用去离子水慢速湿润至土壤中,然后进行18 h以上的闷土处理,达到40%的质量含水率,误差范围控制在3%以内,在闷土期间用保鲜膜包裹土样以减少含水率的变化。将装有原状土的PVC管置于温度可调控的冻融机中,进行不同冻融循环周期的试验。
每年的11月份到次年3月份,黑土一般处于冻结状态;3—5月份,0—20 cm土层内温度变动于0℃上下,表现出“昼融夜冻”的特点[12]。结合拜泉县当地气象资料,设置冻融温度为-10—7℃。该温度下当地冻融周期为20天左右,因此试验设置冻融循环周期为0、1、3、5、7、10、15次和20次。为确保试验过程中土柱可完全冻结、融化,本试验采取的是12 h冻结,12 h融化的缓慢冻融过程。完成冻融循环后将土壤风干过筛,获取5—7 mm团聚体用于结构的测定。
土壤理化性质采用常规方法测定(土壤理化分析1978);土壤机械组成采用吸管法测定;土壤质地划分根据美国制划分标准;土壤容重、饱和持水量、田间持水量和总孔隙度采用环刀法测定;土壤有机质采用重铬酸钾外加热法测定。供试土样相关理化性质如表1所示。
1.2 CT扫描和图像重建
本试验样品图像的获取利用上海光源X射线成像及生物医学应用光束线站(BL13W1)的同步辐射显微CT完成,光子能量设置为30000 eV,分辨率为3.25 μm,曝光时间为1.8 s,样品台与探测器距离为15 cm。将样品固定在样品台上,样品台在水平方向从0到180°匀速旋转,每个样品采集1440张图像。图像重建利用上海光源PITRE软件完成。PITRE软件处理图像首先对图像进行相位恢复,然后将12位投影图像转换为16位,生成正弦图像,然后利用背投影算法重建获取切片图像(图1)。重建后将切片图像存储为8位tiff格式。三维团聚体结构(图1d)的可视化利用Image J完成。

表1 供试土样理化性质Table 1 The physical and chemical properties of studied black soil
总孔隙度=1-容重/比重;砂粒为0.05—2 mm的土壤颗粒、粉粒为0.002—0.05 mm的土壤颗粒、黏粒为<0.002 mm的土壤颗粒

图1 土壤团聚体的投影、正弦、切片和三维结构图像Fig.1 Radiograph,sino,reconstructed slices and 3-D structure of soil aggregate
1.3 图像处理
图像分割是实现数字图像定量分析的必要和关键步骤。由于每个样品包含大量的切片,必须通过批量化处理提高效率。但是不同CT切片图像间亮度差别较大,首先利用Image J软件中的Normalize命令对图像进行归一化处理。对灰度图像的二值分割是土壤结构定量分析的关键,不同分割方法对土壤结构特征分析结果影响很大[13]。为了准确提取土壤孔隙结构数据,比对多种分割模式后采用采用全局阈值法,结合实际的土壤孔隙度反复调试确定每个图像的分割阈值。
1.4 孔隙结构分析
为了避免边界部分的影响,选取团聚体中间部分500×500×500体元进行图像分析。土壤孔隙结构分析利用Image J软件完成。利用Bone J插件计算土壤孔隙的孔隙骨架、分形维数、孔隙连通度。利用3D suit将相连的孔隙从孔隙网络中提取出来后计算孔隙的数量、体积、长度、面积等。按孔隙当量直径将孔隙分为3个等级:非毛管孔隙>100 μm;毛管孔隙30—100 μm;贮存孔隙<30 μm,分别统计出其相应分级孔隙度。孔隙数量是外部孔隙和内部孔隙数量之和。孔隙节点为多个孔隙的连接点。通过公式(1)来计算孔隙形状系数(F)[14]。
F=Ae/A
(1)
式中,Ae为体积与测得孔隙体积相等的球体的表面积,而A为孔隙的实测表面积。按孔隙形状系数将孔隙分为三类:规则型(F≥0.5),不规则型(0.2 分形维数和连通性能够量化不同处理团聚体微结构的差异,很好地反映孔隙网络状况。三维分形维数反映了物体的自相似性和占有空间的有效性,分形维数越大表明该物体结构越复杂,且其值介于2—3之间。孔隙系统的连通性通过计算欧拉特征值(Euler-Poincaré,Ev)来体现,数值越小则孔隙系统连通性越高[9]。本研究中分形维数与欧拉特征值均利用Image J软件获得。 由于试验机时的限制,本研究每组冻融循环采集3个样品进行分析。在数据处理过程中将这3个样品作为3个重复进行分析。利用SPSS 22.0软件进行单因素方差分析,多重比较利用LSD法,显著性水平为0.05。 图2是不同冻融循环次数下团聚体内部结构的二维和三维图像。不同冻融循环次数下团聚体的二维形态有明显的差异(二维图中白色部分为孔隙,黑色部分为固体颗粒)。如图所示,0次冻融循环下的土壤团聚体结构比较致密,随着冻融循环次数的增加,土壤团聚体结构则相对疏松,形成明显的大孔隙结构,中、小孔隙减少。从二维图像可以看出未经冻融时团聚体孔隙以小孔隙为主,随着冻融循环次数的增加孔隙不断增大,5次冻融循环后团聚体出现明显裂隙,孔隙相连,随着冻融循环次数的增多,该现象越为明显。0次冻融循环下的土壤大团聚体内部观察不到中、小团聚体的轮廓和边界。当冻融循环次数达到15次时,团聚体内部孔隙连通呈现网络状,连通的网络状孔隙将大团聚体内部固体颗粒分离,在大团聚体内部可以观察到明显的小团聚体结构。 图2 不同冻融循环次数下土壤团聚体二维和三维结构Fig.2 2D and 3D visualizations of soil aggregate structures under different freeze-thaw cycles 如图2所示,随着冻融循环次数的增加,团聚体孔隙度明显增大。此外,三维图像中黄色部分为立方体边缘孔隙,紫色部分为内部孔隙。从立方体边缘黄色部分的孔隙可以看出,随着冻融循环次数的增加,3次冻融循环后团聚体内部孔隙个体显著增大,单个孔隙在不断增大的同时与相邻孔隙相连通。团聚体内部出现裂隙,孔隙连通度增大,这与二维图像显示结果一致。图像观察表明,多次冻融循环后,团聚体内部孔隙增大,连通性增强,团聚体内部由于孔隙的连通呈现网络状结构。 通过CT 扫描和数字图像处理技术,不仅可以直观可视化研究土壤团聚体的三维结构,还可以定量表征团聚体内部孔隙的连通性和复杂性[17-18]。由于显微CT图像分辨率为3.25 μm,不能分辨出更小的孔隙,所以本文仅研究大于3.25 μm的孔隙。表2为团聚体孔隙的相关参数。 如表2所示,孔隙数量随冻融循环次数的变化并无明显规律。孔隙节点数量随冻融循环次数的增加呈增多趋势,在5次循环以内,孔隙节点数量无规律波动,但达到7次冻融循环以上孔隙节点数量较5次以内明显增大,说明7次以上的冻融循环导致孔隙分支增多。 随着冻融循环次数的增加,土壤团聚体孔隙度不断增大,孔隙度由0次冻融循环后的7.8%增加至20次冻融循环后的23.34%。1次冻融循环后孔隙度增加了7.69%;3次冻融循环后孔隙度增加了77.56%;5次冻融循环后孔隙度增加了96.03%;7次冻融循环后孔隙度增加了92.95%;10次冻融循环后孔隙度增加了142.82%;15次冻融循环后孔隙度增加了206.79%;20次冻融循环后孔隙度增加了199.23%。数据表明,5次冻融循环与7次冻融循环对团聚体孔隙度影响相近;15次冻融循环与20次冻融循环对团聚体孔隙度影响相近。 在7次冻融循环以内,欧拉特征值增减并无规律,7次冻融循环后,欧拉值随冻融循环次数增加而减小,孔隙连通度增大,分形维数值先减小后增大,说明团聚体内部的复杂程度随着冻融循环次数的增加降低而后增大最后趋于平稳。 表2 不同冻融循环次数下土壤团聚体孔隙基本结构参数Table 2 General properties of soil pore network of aggregates under different freeze-thaw cycles 同一行不同小写字母表示差异显著(P<0.05) 图3为不同冻融循环次数下土壤团聚体孔隙大小分布。如图所示,孔径大小分布以>100 μm孔径的非毛管孔隙为主,占总孔隙度60%以上。数据表明,孔径<30 μm的孔隙随着冻融循环次数的增加孔隙度有所减小,相较于0次冻融循环的团聚体,1、3、5、7、10、15次和20次冻融循环后孔隙度分别减小22.22%、7.78%、65.56%、13.33%、62.22%、67.78%和64.44%。孔径在30—100 μm的孔隙随着冻融循环次数的增加孔隙度有所减小,相较于0次冻融循环的团聚体,1、3、5、7、10、15次和20次冻融循环后孔隙度分别减小30.54%、56.16%、68.47%、47.78%、74.88%、78.33%和75.86%。孔径>100 μm的孔隙随着冻融循环次数的增加孔隙度有所增大,相较于0次冻融循环的团聚体,1、3、5、7、10、15次和20次冻融循环后孔隙度分别增加29.42%、149.79%、195.06%、171.81%、272.43%、377.37%和363.58%。随着冻融循环次数的增加,>100 μm孔径的非毛管孔隙孔隙度不断增大;30—100 μm的毛管孔隙孔隙度不断减小;<30 μm的贮存孔隙在7次冻融循环后稳定在总孔隙度的2%以下。 图3 不同冻融循环次数下土壤团聚体孔隙大小分布Fig.3 Pore size distributions of soil aggregates under different freeze-thaw cycles同一孔径分级的不同小写字母表示差异显著(P<0.05) 土壤孔隙形状影响着土壤的水力特性,尤其是瘦长型孔隙,由于其较大的孔壁表面积,更有利于水分和气体的存储[19]。图4表明,瘦长型孔隙随着冻融循环次数的增加孔隙度增大,相较于0次冻融循环的团聚体,1、3、5、7、10、15次和20次冻融循环后孔隙度分别增加17.02%、48.45%、63.20%、53.56%、67.22%、68.56%和69.19%。不规则型孔隙随着冻融循环次数的增加孔隙度有所减小,相较于0次冻融循环的团聚体,1、3、5、7、10、15次和20次冻融循环后孔隙度分别减小23.13%、68.99%、84.71%、72.40%、89.93%、91.28%和93.06%。规则型孔隙随着冻融循环次数的增加孔隙度有所减小,相较于0次冻融循环的团聚体,1、3、5、7、10、15次和20次冻融循环后孔隙度分别减小20.25%、48.53%、78.5%、64.67%、83.94%、86.80%和85.01%。瘦长型孔隙为土壤团聚体孔隙的主要形态,其所占孔隙度大约为60%,且经过冻融处理后瘦长型孔隙度不断提高后趋于稳定,20次冻融循环后瘦长型孔隙占总孔隙度96%。而规则型和不规则孔隙所占孔隙度比例呈现出相反的趋势。这与图1中观察到的现象一致。随着冻融循环次数的增加,土壤团聚体中产生了更多的细长的不规则的孔隙,呈现明显的复杂多孔结构。 图4 不同冻融循环次数下土壤团聚体孔隙形状分布Fig.4 Pore shape distribution of soil aggregates under different freeze-thaw cycles同一孔隙形状的不同小写字母表示差异显著(P<0.05);各型孔隙体积占总孔隙体积的百分比为规则型孔隙度,不规则型孔隙度和瘦长型孔隙度 土体的冻融过程,实质上就是土中水的冻结和融化过程,当土中水冻结时,体积膨胀,使得土颗粒间的孔隙体积增大,而当冰融化时,孔隙体积减小,所以冻融过程将会改变土颗粒间的结构联接,排列方式,从而改变土壤结构[20-21]。此外,冻融作用会造成土颗粒之间原始固有胶结逐渐减弱,黏聚力不断降低[22]。 孔隙度是受冻融循环影响最基础、直观的孔隙特征参数。冻融初期,未经过冻融循环的团聚体孔隙内含有一定水分。冻融循环开始时,孔隙中的水转化成冰,体积增大,导致团聚体孔隙变大;冰晶融化时,孔隙未能恢复冻结前状态,此过程循环往复。此外,随着冻融循环次数的增加,被冰晶挤压而产生形变的孔隙恢复能力逐渐减弱。以上两个因素导致孔隙体积不断增大,团聚体孔隙度随冻融循环次数的增多持续增长状态。当冻融循环达到15次以后原有的结构已经被改变,团聚体内部裂隙明显,在大团聚体内部形成多个小团聚体,达到新的稳定状态,受冻融循环的影响减小甚至不再受其影响。但在试验过程中发现此时的大团聚体结构脆弱,极易破碎。多次的冻融循环使得土壤孔隙结构不断变化,但当冻融循环次数在15次以上时孔隙结构变化不再明显。 在冻融循环过程中孔隙数量并未随冻融循环次数增加呈现规律性变化。该现象是由于在水与冰的相变过程中除孔隙体积的增长外也会有新的孔隙形成,因此孔隙数量随冻融循环次数的增加并无规律性变化;从切片分析结果来看,内部孔隙数量与外部孔隙数量变化也无规律性变化。而孔隙节点数量增加是由于冻融循环导致孔隙出现裂隙或者多个孔隙由于不断扩大相连,这与瘦长型孔隙孔隙度占比不断增大原理相同。 连通性与分形维数能很好的反应团聚体形态结构的变化[4]。欧拉指数判定孔隙连通性结果表明,冻融循环次数达到7次以上有改善团聚体孔隙连通性的作用,而分形维数并未受冻融循环影响呈现规律性变化。 受冻融作用的影响,非毛管孔隙、毛管孔隙和贮存孔隙的孔隙度变化显著。冻融作用对土壤结构的改变在不同冻融循环次数下有所不同,由于该过程对团聚体内部结构的改变存在着复杂的不确定性,因此在3、5次和7次冻融循环下孔隙分级的规律性略微不同,但7次冻融循环后规律清晰,整体趋势显著。非毛管孔隙度比例随冻融循环次数增加不断增大,而毛管孔隙与贮存孔隙占比不断减小,该现象是由于冻融作用的循环往复导致小孔隙体积增大,促进了大孔隙的形成,进而使非毛管孔隙、毛管孔隙和贮存孔隙的占比产生变化。而土壤团聚体大孔隙的形成,进一步导致水分运移速率增加。孔径较大的大孔隙具有较强的水力传导性,但这种促进作用是有限的,孔隙之间的连通性也是决定水流运移速率的主要因素[23]。因此,冻融循环对水分运移速率的增加产生一定促进作用。 冻融循环对孔隙形状可产生影响。结合图2中二维、三维图像与图4的定性、定量分析表明,随着冻融循环次数的增加,规则孔隙与不规则孔隙的孔隙度占比均减小,而瘦长型孔隙占比显著增大,这与冻融循环造成大团聚体内部产生裂隙直接相关。此外,原本的单个孔隙相连通也使得孔隙形状更趋近于瘦长型,造成规则与不规则孔隙的占比减小。由于瘦长型孔隙的增多更有利于水分与气体的存储[20],因此,冻融循环对土壤水分与气体的存储和运移产生一定影响。 本文利用同步辐射显微CT和数字图像处理与分析技术研究团聚体三维微结构,实现了团聚体内部孔隙结构的可视化。结果表明,随着冻融循环次数的增加,团聚体孔隙度增大,在15次冻融循环后趋于稳定;瘦长型孔隙孔隙度占比增加,规则与不规则孔隙趋势相反;>100 μm孔隙占比不断增大;孔隙连通性在7次冻融循环后显著改善;孔隙节点数量随冻融循环次数的增加而增大;孔隙数量和分形维数无显著性变化。该研究为冻融侵蚀的研究以及春季解冻期水土流失的防治提供理论依据。此外,冻融循环对黑土团聚体孔隙特征产生影响的同时,对土壤水分的存储与运移以及团聚体破碎机制产生一定影响,而国内外关于该方面影响的研究尚且不足。1.5 统计分析
2 结果与分析
2.1 土壤团聚体可视化
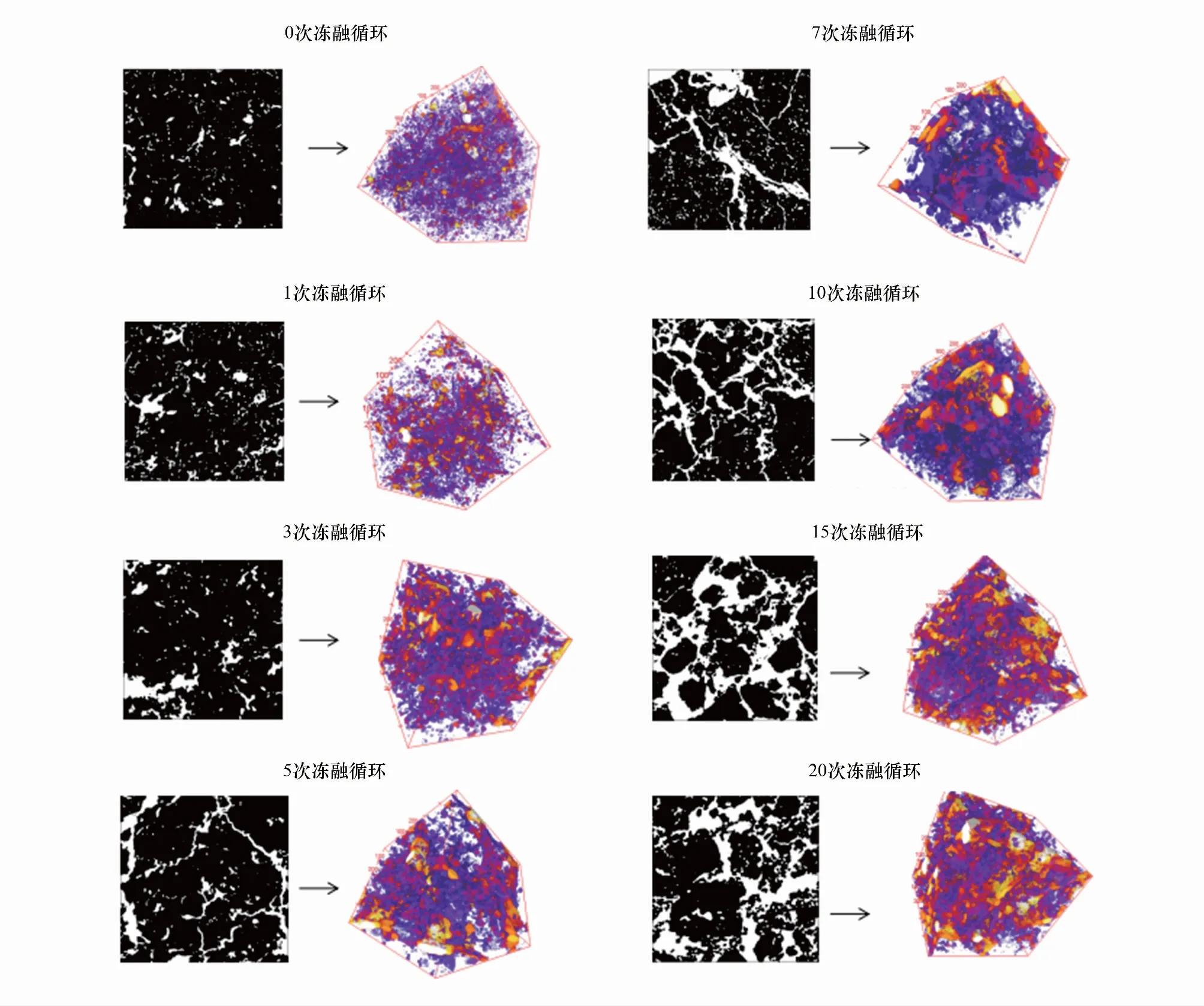
2.2 团聚体孔隙基本特征

2.3 团聚体孔隙分布特征

2.4 土壤团聚体孔隙形状特征

3 讨论
4 结论
