异质生境对生防昆虫莲草直胸跳甲数量分布的影响
2019-06-27吴昊代文魁蔡水清
吴昊 代文魁 蔡水清

摘要 昆虫的种群动态和数量分布与环境因素密切相关。本研究通过在中国南方空心莲子草大规模入侵的区域分别设置水、陆样地,采用曲线回归和冗余分析等方法探讨了异质生境中空心莲子草形态特征、入侵群落植物多样性及无机环境对生防昆虫莲草直胸跳甲数量分布的影响。结果显示:水、陆两境中莲草直胸跳甲数量均随着空心莲子草株高增加而上升;莲草直胸跳甲数量随水生型空心莲子草分枝数及入侵盖度的增加而上升。莲草直胸跳甲数量与陆生群落植物物种多样性无显著相关性,但在水生环境中,莲草直胸跳甲数量随群落的Simpson、Shannon和Pielou指数上升而显著下降。水、陆两境中决定莲草直胸跳甲数量的主导性环境因子均为年均气温、纬度和海拔。研究结果可为入侵植物的生物防治及引进天敌提供参考。
关键词 环境因子; 莲草直胸跳甲; 种群数量; 生物入侵; 冗余分析
中图分类号: S 476
文献标识码: ADOI: 10.16688/j.zwbh.2018246
Abstract The insect population dynamics and quantity distribution are closely related to environmental factors. In this study, we set up aquatic and terrestrial plots in locations that were invaded largely by alien Alternanthera philoxeroides in Southern China, by using the curvilinear regression and redundancy analysis methods; we analyzed the influences of A.philoxeroides morphological characteristics, plant species diversity within invaded community and inorganic environmental factors on the population abundance of an biocontrol insect Agasicles hygrophila in heterogeneous habitats. The results showed that the A. hygrophila abundance increased with the increasing A.philoxeroides height in both aquatic and terrestrial habitats, and increased with A.philoxeroides branches and invasion coverage only in the aquatic habitat. The A.hygrophila abundance had no significant relationships with the terrestrial plant species diversity; however, it significantly decreased with the rising Simpson, Shannon, Pielou indices in aquatic invaded plant community. In addition, the dominant environmental factors which determined the A.hygrophila abundance were annual mean temperature, latitude and elevation in both aquatic and terrestrial habitats. Our research could provide suggestive references for biological control of invasive plants and introduction of natural enemies.
Key words environmental factor; Agasicles hygrophila; population abundance; biological invasion; RDA
昆蟲作为自然界数量最多的动物占据着各类微生境,其对环境变化敏感,昆虫种群的分布与环境关系极为紧密[13];分析昆虫生境因子(生物、非生物)耦合关系对于研究昆虫、防治昆虫或培养利用昆虫具有重要指导意义[4]。以往研究表明,水温、总磷、氨态氮、海拔和溶解氧等环境驱动因子在不同程度上影响水生昆虫群落结构及其多样性[56]。同时,昆虫分布状况也受到生境中植物群落特征的影响,如:在长白山阔叶红松林中,植食性昆虫丰富度与植物群落物种丰富度呈负相关,植食性昆虫类群主要依靠草本的复杂性和多样性[7];而在油松林中,随着植物群落科种组成复杂程度的增加,昆虫群落的多样性增加[8]。此外,郭海滨等关于草坪昆虫群落的研究表明,植食性害虫的分布与杂草种类及草坪高度呈负向关系[9]。在全球环境迅速变化的背景下,探讨昆虫(尤其是天敌昆虫)的数量分布规律及其关键性影响因子,有助于维护农业生态系统稳定性及保障国家生物安全[10]。
莲草直胸跳甲Agasicles hygrophila又名喜旱莲子草叶甲、曲纹叶甲,属鞘翅目叶甲科跳甲亚科。其起源于南美洲,取食空心莲子草Alternanthera philoxeroides叶片及上部嫩叶茎,幼虫钻入茎秆后化蛹并分泌有毒物质阻止节间生长。莲草直胸跳甲于1986 年从美国佛罗里达引入中国,是防治入侵植物空心莲子草最有效的专食性天敌昆虫[11]。近期,在宏观水平上关于莲草直胸跳甲的研究主要集中于生物学特征、繁殖技术及其生防效果评估等方面,如:前人研究表明,莲草直胸跳甲在旱地的扩散能力强于湿地[12];适当延长光周期的光照时数可提高莲草直胸跳甲幼虫的存活率[13];在有限的空间及资源条件下,每头莲草直胸跳甲成虫所需的最适空间约为31.25 cm3[14];生境中的养分富集会导致空心莲子草对莲草直胸跳甲取食的诱导抗性增加,从而降低莲草直胸跳甲的生防效应[15]。
尽管诸多研究已表明环境异质性是决定群落结构和生态系统健康的重要因素[16],但迄今为止,关于大尺度地理格局上“环境因子莲草直胸跳甲种群分布”的研究尚未见报道。本研究通过大尺度的野外调查,对比分析了水生及陆生型空心莲子草入侵群落中莲草直胸跳甲数量的差异,并分别从无机生境因子、入侵群落植物多样性、空心莲子草种群特征等方面探讨其对莲草直胸跳甲数量的影响,以期阐明莲草直胸跳甲在异质生境中的分布规律及其主导性因素,为入侵植物的生物防治及评估天敌昆虫的生防效应提供依据。
1 材料与方法
1.1 野外调查
2014年7-8月,在中国南方21°N~31°N纬度范围内,选择空心莲子草入侵面积>100 m2的区域设置调查样地。每隔2个纬度梯度设置4~6个陆生样地,并在同纬度段生境状况较为相似的地点设置4~6个水生样地。共计布设样地52个,分布于中南5省12市,其中陆生29个,水生23个(表1)。研究样地处于中国亚热带季风气候区,水热条件优越,年平均气温20℃,年均降水量1 348 mm,土壤质地以赤红壤和黄壤为主,多呈酸性。空心莲子草入侵生境包括撂荒地、田埂、河岸带、水库、池塘和水田等多种类型。
选择晴朗无风的天气进行野外调查。每个样地的面积为10 m×10 m,在样地中央均匀设置3条10 m长的样带,每条样带均匀布设5个0.5 m×0.5 m的小样方,调查时记录样方中所有植物的物种名称、高度、盖度、多度。对于未知植物,拍照或采标本带回实验室后进行物种鉴定。植物调查结束后,利用伸缩式捕虫网(直径30 cm,网深60 cm)沿着每个样地中任意一条样带来回横扫两次,将搜集到的昆虫全部装入昆虫瓶中,统计不同生境空心莲子草群落中单位面积的莲草直胸跳甲种群数量。
1.2 环境测定
利用手持式GPS定位仪(Garmin eTrex20)记录样地生境的经度、纬度和海拔。在所有调查结束后,对于陆生样地,沿着样带均匀设置5个土壤采集点,共采集土壤约250 g,装入塑封袋置于车载冰箱(MobicoolCF50DC)中冷冻保存,带回实验室后利用全自动化学分析仪(Easychem Plus)测定土壤中的氨態氮、硝态氮等肥力指标。对于水生样地,直接使用YSI便携式水质分析仪(Professional Plus)测定水体中氨态氮、硝态氮、溶解氧和电导率等水质指标。在中央气象台国家气象中心网站(http:∥www.nmc.gov.cn/)查询获取各个调查地点近50年来的年均气温及年均降雨量数据。
1.3 数据计算
1.3.1 植物重要值
重要值(importance value, IV)是度量群落中植物生长及相对优势度的综合指标,各植物重要值计算公式如下[17]:IV=(相对盖度+相对高度+相对多度)/3。
1.3.2 植物群落α物种多样性指数
Patrick 丰富度指数:R=S;Simpson多样性指数:λ=1-ΣP2i;ShannonWiener 多样性指数:H=-ΣPi× lnPi;Pielou 均匀度指数:E=H/lnS[18]。式中:S为样地群落内所有植物的物种数目,Pi 为第 i 种植物的相对重要值。
1.4 统计分析
利用SPSS 16.0数理统计软件分别对水陆两境中空心莲子草形态特征(株高、分枝数、入侵盖度)、植物群落4个α物种多样性指数与样地中单位面积莲草直胸跳甲的数量进行回归分析。以SPSS 16.0提供的11种曲线方程建立回归模型并检验其显著性,选择回归显著(P<0.05)且拟合系数最高的模型进行分析。建立环境因子矩阵(经度、纬度、海拔、氨态氮、硝态氮、年均温、年均降雨量,52×7)和莲草直胸跳甲数量矩阵(52×1),利用数量生态学软件CANOCO 4.5提供的冗余分析(redundancy analysis,RDA)探讨多环境变量对异质生境中莲草直胸跳甲分布的影响。利用SPSS 16.0分析单环境变量与莲草直胸跳甲数量之间的回归拟合关系,并对水、陆两境的莲草直胸跳甲数量进行独立样本t检验(independentsample t test)。
2 结果与分析
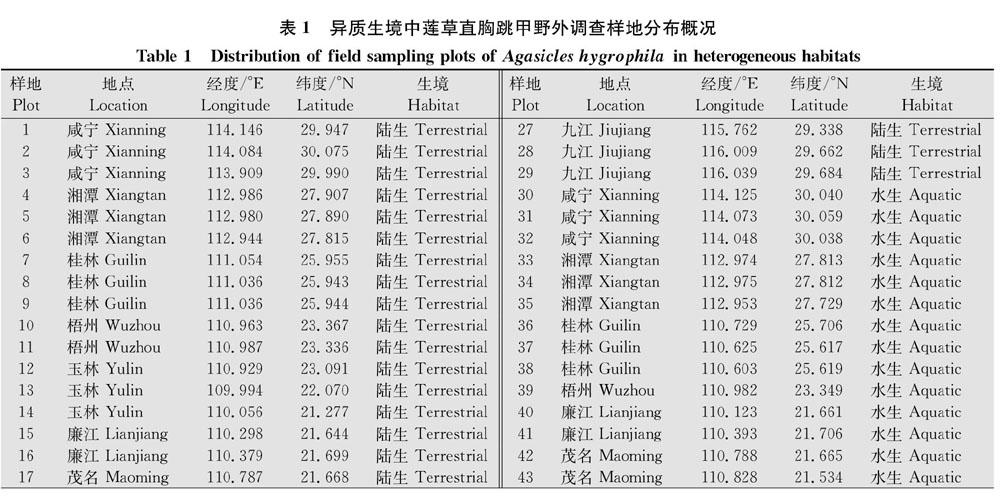
2.1 异质生境中空心莲子草形态特征对莲草直胸跳甲数量的影响
对陆生生境中空心莲子草的3项形态特征(株高、无性分枝数、入侵盖度)与单位面积内的莲草直胸跳甲数量进行回归分析,结果表明:空心莲子草株高与莲草直胸跳甲数量之间具有显著的二项式回归关系(P=0.001),随着空心莲子草株高增加,莲草直胸跳甲数量呈上升趋势;尤其当空心莲子草株高>30 cm时,莲草直胸跳甲数量的上升趋势更为明显(图1a)。陆生生境中空心莲子草的无性分枝数、入侵盖度与莲草直胸跳甲数量均无显著相关性。
水生生境的空心莲子草无性分枝数、株高、入侵盖度与单位面积的莲草直胸跳甲数量之间均呈显著的回归关系(图1)。其中,空心莲子草无性分枝数(P=0.038)、入侵盖度(P=0.048)与莲草直胸跳甲数量呈显著线性回归关系,随着空心莲子草分枝数及盖度上升,莲草直胸跳甲数量呈明显上升趋势(图1b、d)。空心莲子草株高(P=0.002)与莲草直胸跳甲数量呈显著二项式回归关系,与陆生生境相似,当水生空心莲子草株高>30 cm时,莲草直胸跳甲数量急剧上升(图1c)。
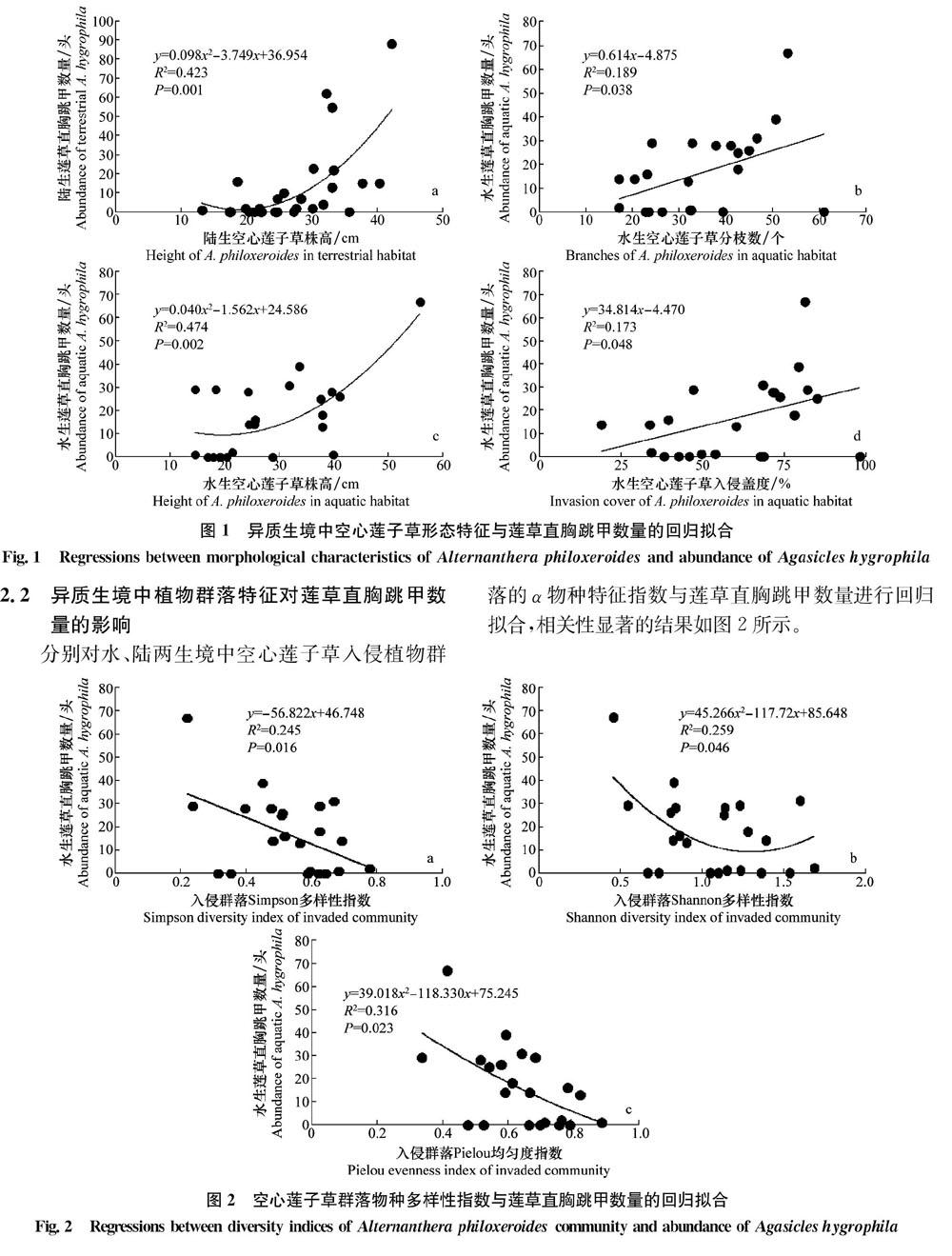
2.2 异质生境中植物群落特征对莲草直胸跳甲数量的影响
分别对水、陆两生境中空心莲子草入侵植物群落的α物种特征指数与莲草直胸跳甲数量进行回归拟合,相关性显著的结果如图2所示。
2.3 异质性环境因子对莲草直胸跳甲数量的影响
RDA是研究生态系统中物种沿着环境梯度分布格局的常用数量排序方法,其属于约束性模型,可直观反映出多个环境变量对生物的综合效应。通过对水、陆两境中莲草直胸跳甲数量与各个环境因子进行RDA排序,发现两类生境中RDA前两排序轴均包含了100%的生态信息(陆生生境:RDA 1轴 55.1%,RDA 2轴 45.9%;水生生境:RDA 1轴 63.7%,RDA 2轴 46.3%),表明莲草直胸跳甲数量很大程度上受到所测环境因子的影响,故采用前两个排序轴作RDA二维排序图(图3)。其中,RDA第1轴对莲草直胸跳甲数量起主导性作用,环境因子与RDA 1轴相关系数的分析结果如表2所示。
对于陆生生境而言,RDA 1轴中决定莲草直胸跳甲数量的主导性环境因子为年均气温(相关系数-0.822,P<0.001)和纬度(相关系数0.716,P<0.001)。入侵群落中莲草直胸跳甲数量与年均气温呈显著的负向关系,而与纬度呈密切的正向关系。对于水生生境而言,RDA 1轴中决定莲草直胸跳甲数量的主导性环境因子分别为年均气温(相关系数-0.897,P<0.001)、纬度(相关系数0.817,P<0.001)和海拔(相关系数0.571,P<0.001);其中,莲草直胸跳甲数量与年均气温呈显著的负向关系,而与纬度、海拔呈较大的正向关系。经度、氨态氮、硝态氮、年均降雨量均未对异质生境中莲草直胸跳甲的数量造成显著性影响。
为了更加精细地探讨莲草直胸跳甲数量与各环境因子之间的逐一对应关系,对莲草直胸跳甲数量环境因子进行单因素回归分析,结果如图4所示。水生生境中,纬度(拟合系数0.176,P=0.047)、海拔(拟合系数0.238,P=0.018)均与莲草直胸跳甲数量之间存在显著的正向直线拟合关系,随着纬度和海拔的上升,莲草直胸跳甲的数量显著增加;年均气温(拟合系数0.246,P=0.016)与莲草直胸跳甲数量之间存在显著的负向直线拟合关系,莲草直胸跳甲数量随着气温的上升而急剧下降。陆生生境中,年均气温(拟合系数0.191,P=0.018)也与莲草直胸跳甲数量之间存在显著的负向直线拟合关系,随着气温升高,莲草直胸跳甲数量明显下降。以上单因素回归分析的结果与RDA排序(图3)基本吻合,这表明:无论是多环境变量综合作用还是单环境因素独立影响,对莲草直胸跳甲种群数量起决定性影响作用的环境因子均为气温、纬度和海拔。但与RDA排序不同的是,在回归分析中,年均降雨量与陆生生境的莲草直胸跳甲数量之间具有显著的二项式拟合关系(拟合系数0.260,P=0.020)(图4e),随着降雨量增加,莲草直胸跳甲数量呈现“先下降后上升”的曲线变化趋势。此外,独立样本t检验表明水、陆两境的莲草直胸跳甲数量并无显著性差异(t=-0.874, P=0.386)。
3 结论与讨论
植物和昆蟲是生态系统的重要组分,二者互作关系及其演化深刻地影响生态系统结构和功能[19];由于植物所处环境存在较大的异质性,同种植物的不同生态型也会对其天敌昆虫的生长发育及种群动态造成显著影响[20]。入侵植物空心莲子草具备强大的表型可塑性,而莲草直胸跳甲在空心莲子草茎秆中钻蛀化蛹是其完成生活史的关键环节。相关研究表明,水生型空心莲子草茎秆的髓部薄壁细胞较薄、髓腔大,莲草直胸跳甲化蛹率高;而陆生型空心莲子草髓腔基本为实心、维管柱厚,莲草直胸跳甲化蛹能力较弱[11,21]。此外,陆生环境中空心莲子草体内的单宁、总酚等防御性次级代谢物质含量高于水生环境,能够对莲草直胸跳甲取食产生较强的化学防御能力[22];故相较于陆生型空心莲子草而言,莲草直胸跳甲数量与水生型空心莲子草形态特征呈现较大的正向拟合关系(图1)。马瑞燕等研究发现莲草直胸跳甲化蛹率与茎秆节间长呈正相关[21],本研究中,水、陆两境空心莲子草株高均与莲草直胸跳甲数量呈显著正向关系(图1a, c),可能是由于植株越高,则节间距相应越长,能够为跳甲产卵及幼虫孵化提供更多空间。
植物群落物种多样性指数反映了群落内植物种类数目及各物种在种间分布的均匀程度[23],植物多样性的增加一定程度上能够抑制入侵植物在群落中的优势度[24]。对于水生型空心莲子草群落而言,一方面,其群落结构相对简单,且莲草直胸跳甲对水生型空心莲子草的取食专一性及生防成功率较高[2526],而其他水生植物的存在抑制了空心莲子草数量,即相应降低了莲草直胸跳甲的产卵繁殖场所及其食物来源;另一方面,诸多研究表明,较高的植物多样性能够支持更加多样化的昆虫群落[8,2728],进而可能对莲草直胸跳甲生长繁殖及种群扩增产生竞争抑制作用,故莲草直胸跳甲数量与水生型空心莲子草入侵群落的Simpson、Shannon及Pielou物种多样性指数呈显著负向关系(图2)。但本研究发现陆生植物多样性并未对莲草直胸跳甲数量产生显著影响,可能是因为陆生环境相对于水生环境的条件更为复杂,空心莲子草在陆生生境中面对更多的取食者[8],导致植物多样性与莲草直胸跳甲数量之间的关系不明显。
环境温度是制约变温动物(尤其是昆虫)种群增长的重要因素[29]。莲草直胸跳甲生长发育需要一定的有效积温,但年均气温若过高,则容易造成昆虫出现各种热害反应,如:体内水分散失加快、产卵量降低、发育受阻等,最终导致其种群数量急剧下降[30]。纬度包含复杂的环境因子,如:温度、生长季长度、光周期、土壤湿度等,纬度梯度造成的水热条件非均衡性导致入侵植物群落结构显著改变[31]。随着纬度上升气温逐渐降低,Wu等发现中国南方21°N~31°N范围内,强大的生态适应性使得空心莲子草在植物群落中的入侵优势度随纬度上升而被加剧[32],这可能为莲草直胸跳甲的生长繁殖提供了更多的食物来源及栖息地,导致莲草直胸跳甲数量与纬度呈显著正相关。海拔变化影响植物的资源分配策略,前人研究表明,高海拔地区植物的叶片寿命更长、植物种群为了获取更多水热资源会相应增加其地上生物量[3334],有利于维持更多的天敌昆虫数量;此外,高海拔地区人类活动干扰较小、水流较缓、可减少对水生生境中莲草直胸跳甲卵块的冲刷[3536],这可能是水生环境中莲草直胸跳甲数量与海拔呈正相关的主要原因。
生境是昆虫生存的重要因素,不同生境格局显著影响昆虫的繁殖、分布与扩散[37],在全球环境迅速变化的背景下,天敌昆虫对外来植物入侵响应生境变化及其群落动态结构具重要的调控作用[38]。本研究综合分析了异质性生境中入侵植物形态特征、入侵生境的植物群落特征以及无机环境因子对生防昆虫莲草直胸跳甲数量分布的影响,可为预测环境变化下外来植物入侵态势及天敌昆虫引种评价等提供参考。本研究中降雨量与陆生生境的莲草直胸跳甲数量形成显著的“先下降后上升”回归拟合关系,这与RDA排序结果不相一致,可能是由于RDA中多环境因子之间存在交互效应(如:降雨量氮素)或共变效应(如:经度降雨量)。关于降雨量、氮素等因素对莲草直胸跳甲的影响尚需进一步设置人工控制试验加以深入探讨。
参考文献
[1] 康乐.环境胁迫下的昆虫-植物相互关系[J].生态学杂志,1995,14(5):5157.
[2] 杜瑞卿,王庆林,张征田,等.EPT昆虫群落分布与环境因子的相关性[J].昆虫学报,2008,51(3):336341.
[3] 聂传朋,李焰焰,李永民.人工草坪昆虫多样性与环境因子的相关性初步研究以庐山风景区为例[J].长江流域资源与环境,2014,23(5):729734.
[4] BRNDLE M, AMARELL U, AUGE H, et al. Plant and insect diversity along a pollution gradient: understanding species richness across trophic levels [J]. Biodiversity & Conservation, 2001, 10(9): 14971511.
[5] 秦春燕,张勇,于海燕,等.不同类群水生昆虫群落间的一致性以及空间和环境因子的相对作用[J].生物多样性,2013,21(3):326333.
[6] 刘曼红,李亚俊,张宇,等.帽儿山溪流水生昆虫与环境因子的相关性[J].吉林农业大学学报,2014,36(4):479487.
[7] 董百丽,姬兰柱,魏春艳,等.长白山阔叶红松林植物群落与昆虫群落的相互关系研究[J].生态学杂志,2005,24(9):10131016.
[8] 高宝嘉,张执中,李镇宇.植物群落对昆虫群落及种群生态效应的数学分析[J].生态学报,1993,13(2):130134.
[9] 郭海滨,李保平,强胜,等.狗牙根草坪昆虫群落组成与环境因子相关性研究[J].南京农业大学学报,2009,32(3):6370.
[10]张礼生,陈红印.我国天敌昆虫与生防微生物资源引种三十年成就与展望[J].植物保护,2016,42(5):2432.
[11]马瑞燕,丁建清,李佰铜,等.莲草直胸跳甲在不同生态型空心莲子草上的化蛹适应性[J].中国生物防治,2003,19(2):5458.
[12]李彦宁,傅建炜,郭建英,等.莲草直胸跳甲释放量对其种群构建的影响[J].生物安全学报,2011,20(4):275280.
[13]赵龙龙,刘蓉,郭艳琼,等.光周期对莲草直胸跳甲幼虫发育的影响[J].山西农业大学学报(自然科学版),2014,34(3):193197.
[14]冀佳悦,赵龙龙,贾少华,等.虫口密度对莲草直胸跳甲生殖力和存活能力的影响[J].山西农业大学学报(自然科学版),2014,34(5):400402.
[15]周方,张致杰,刘木,等.养分影响入侵种喜旱莲子草对专食性天敌的防御[J].生物多样性,2017,25(12):12761284.
[16]WARREN D S, ALT M, OLSON D K, et al. The relationship between the spectral diversity of satellite imagery, habitat heterogeneity, and plant species richness [J].Ecological Informatics, 2014, 24: 160168.
[17]JING Zhaobin, CHENG Jimin, SU Jishuai, et al. Changes in plant community composition and soil properties under3decade grazing exclusion in semiarid grassland[J]. Ecological Engineering, 2014, 64: 171178.
[18]WU Hao, CARRILLO J, DING Jianqing. Species diversity and environmental determinants of aquatic and terrestrial communities invaded by Alternanthera philoxeroides[J]. Science of the Total Environment, 2017, 581582: 666675.
[19]ERWIN A C, ZST T, ALI J G, et al. Aboveground herbivory by red milkweed beetles facilitates aboveand belowground conspecific insects and reduces fruit production in common milkweed [J]. Journal of Ecology, 2014, 102: 10381047.
[20]LUO Fangli, CHEN Yue, HUANG Lin, et al. Shifting effects of physiological integration on performance of a clonal plant during submergence and desubmergence [J]. Annals of Botany, 2014, 113(7): 12651274.
[21]马瑞燕,王韧.不同生态型的喜旱莲子草对莲草直胸跳甲化蛹能力的影响[J].植物生态学报,2004,28(1):2430.
[22]高芳磊,郭素民,闫明,等.不同生境下空心莲子草响应模拟昆虫采食的生长和化学防御策略[J].生態学报,2018,38(7):23442352.
[23]CADOTTE M W, CARSCADDEN K, MIROTCHNICK N. Beyond species: Functional diversity and the maintenance of ecological processes and services [J]. Journal of Applied Ecology, 2011, 48: 10791087.
[24]吴昊,丁建清.入侵生态学最新研究动态[J].科学通报,2014,59(6):438448.
[25]马瑞燕,王韧.喜旱莲子草在中国的入侵机理及其生物防治[J].应用与环境生物学报,2005,11(2):246250.
[26]吴昊.纬度梯度及生境异质性对空心莲子草入侵效应的影响[D].武汉:中国科学院研究生院(武汉植物园),2015.
[27]朱慧,彭媛媛,王德利.植物对昆虫多样性的影响[J].生态学杂志,2008,27(12):22152221.
[28]魏永平,张雅林,汪晓光.黄土高原苹果园植物多样性及其对昆虫群落的影响[J].西北植物学报,2010,30(12):25172523.
[29]韩宗礼,谭晓玲,陈巨莲.环境温度变化对异色瓢虫的飞行与运动能力的影响[J].中国生物防治学报,2017,33(4):433441.
[30]李建宇,史梦竹,郑丽祯,等.温度对2个地理种群莲草直胸跳甲成虫产卵量及存活率的影响[J].生物安全学报,2013,22(1):5760.
[31]吴昊.入侵植物空心莲子草春季沿纬度变化的群落特征[J].浙江农林大学学报,2017,34(5):816824.
[32]WU Hao, CARRILLO Juli, DING Jianqing. Invasion by alligator weed, Alternanthera philoxeroides, is associated with decreased species diversity across the latitudinal gradient in China [J]. Journal of Plant Ecology, 2016, 9(3): 311319.
[33]宋璐璐,樊江文,吴绍洪.植物叶片性状沿海拔梯度变化研究进展[J].地理科学进展,2011,30(11):14311439.
[34]张茜,赵成章,董小刚,等.高寒退化草地不同海拔狼毒种群花大小、数量与个体大小的关系[J].生态学杂志,2013,32(12):31603166.
[35]李强,杨莲芳,吴璟,等.西苕溪EPT昆虫群落分布与环境因子的典范对应分析[J].生态学报,2006,26(11):38173825.
[36]傅建炜,马明勇,郭建英,等.水流对莲草直胸跳甲种群存活和扩散的影响[J].生物安全学报,2011,20(4):281284.
[37]张金钰,李鑫,吴素蓉,等.果园生境对金纹细蛾寄生蜂发生影响及优势寄生蜂空间分布研究[J].中国生物防治学报,2012,28(3):326333.
[38]LU Xinming, SIEMANN Evan, HE Minyan, et al. Warming benefits a native species competing with an invasive congener in the presence of a biocontrol beetle [J]. New Phytologist, 2016, 211(4): 13711381.
(責任编辑:田 喆)
