地形对喀斯特山地植物物种多样性及分布格局的影响
2019-06-14李芹,容丽,王敏
李 芹, 容 丽, 王 敏
(贵州师范大学 地理与环境科学学院, 贵州 贵阳 550001)
物种多样性是生物多样性在物种水平上的表现形式,对物种多样性的研究一直是生态学科研究的重要内容[1]。地形是影响区域群落和物种变化的主要环境因子之一,通过海拔高度、坡位、坡度、坡向等地形因子的改变引起光照、温度、湿度、土壤水分、养分等环境因子的改变[2],进而影响植物群落组成、分布及物种多样性。地形差异形成的生境异质也为群落中多物种的共存提供了条件,有利于生物多样性的维持[3-4]。植物与地形在不同尺度上都有着密切的联系,研究两者间的关系可以让我们更好地了解生态系统功能的形成及发展过程,对保护生物的多样性具有重要意义[5]。长期以来,学者们基于地形因素对群落物种多样性与分布格局的形成及其维持机制进行了大量探索,取得了不少重要发现。海拔被认为是影响物种多样性分布格局的决定性因素之一[6],有研究[7]表明随着海拔的上升,物种的丰富度和密度呈现先升高后下降的趋势,也有研究[8]表明随着海拔的升高物种多样性呈现不同的分布格局,关于海拔对物种多样性分布格局的影响机制并未得到普遍共识。此外,坡位、坡向和坡度对物种多样性和分布都具有不同程度的影响[9-10]。
喀斯特地区因其特殊的岩溶构造,发育了独特的地上地下二元空间结构,地表储水困难,生境具有石漠化、土壤贫瘠、含钙量高等特点,植被也因此具有旱生性、岩生性等不同于其他地区植被的特点。生境与植被的独特性也吸引了众多学者的研究,但大多集中在植被群落动态演替[11]、生境特征[12]、生态恢复[13]、物种组成与多样性特征[14-15]等方面,在地形对物种多样性的影响方面的研究尚不多见。此次研究将探讨白云岩喀斯特地区地形对物种多样性的影响。黔东南施秉县境内分布有大面积的白云岩喀斯特,在地形地貌的发育和植被分布上都与石灰岩喀斯特存在差异,具有其独特的地质地貌条件。施秉白云岩喀斯特发育并保存了大面积的原生森林,物种组成丰富多样。据调查,区内野生种子植物共有140科538属1 087种,其中包括被子植物133科521属1 061种,裸子植物7科17属26种[16-17]。从地理成分上看,科的区系分布较多为热带分布(42.86%),世界分布(32.14%),其次是温带分布(25%);属的分布类型中温带分布占50.82%,热带分布占46.91%,中国特有属占2.26%。总体上看,植物区系主要以热带分布和温带分布占主体[17],是中国南方亚热带喀斯特原生森林保存较完好,物种种类比较复杂多样的地区之一,也是白云岩喀斯特地貌比较典型的代表。本文以贵州省施秉县云台山白云岩喀斯特世界自然遗产地为研究对象,比较不同地形条件下各生长型植物的物种多样性差异,并对物种与地形因子进行排序分析,探讨地形对喀斯特区物种多样性及其空间分布的影响,为喀斯特地区的生态恢复及生物多样性保护提供科学依据。
1 材料与方法
1.1 研究区基本概况
研究区为世界遗产地的施秉白云岩喀斯特,经度108°01′34″—108°09′32″E,纬度27°05′49″—27°13′59″N间,边界与云台山景区—杉木河景区—生态水源涵养区重合,总面积28 295 hm2,核心区面积为10 280 hm2,缓冲区面积为18 015 hm2。地势由西、西北向东、东南逐渐降低,最高海拔1 869 m,最低海拔486 m,平均海拔526 m。气候属于亚热带季风湿润气候区,年平均气温为16 ℃,降水丰沛,年平均降水量1 220 mm,集中分布在4—10月,气候温和湿润,四季如春,具有亚热带山地湿润气候的特点。成土母岩为寒武纪纯白云岩,土壤为白云岩风化的薄层石灰土。
1.2 地貌发育特征
施秉白云岩喀斯特发育于寒武纪白云岩上,具有比较完整而典型的白云岩喀斯特地貌。与石灰岩喀斯特地区不同的是:石灰岩更易溶,侵蚀以化学溶蚀为主,地上地下喀斯特发育强烈;而白云岩相对不易可溶,喀斯特作用不及石灰岩地区强烈,地貌发育以物理崩塌为主[18]。施秉白云岩喀斯特地貌发育以河流下切侵蚀和崩塌作用为主,景观上呈现出峰丛峡谷和峰丛谷地等地貌景观,山体险峻陡立,只有局部地区发育有峰丛浅洼和塔状孤峰,在景观表现和地貌发育上都与石灰岩喀斯特不同。
1.3 样地设置与调查
采用典型样方法,在遗产地核心区的不同地形部位设置样地。根据山体划分山顶、上坡、中坡、下坡、山脊和河谷几个坡位;坡向分阴坡(337.5°~22.5°,22.5°~67.5°)、阳坡(157.5°~202.5°,202.5°~247.5°)、半阴坡(67.5°~112.5°,292.5°~337.5°)、半阳坡(112.5°~157.5°,247.5°~292.5°);坡度分平坡(<5°)、缓坡(6°~15°)、斜坡(16°~25°)和陡坡(>25°)4个等级。根据地形和植被组成等情况,在每个样地内设置10 m×10 m或20 m×20 m的乔木样方若干。在每个20 m×20 m的乔木样方对角线处各设置5 m×5 m的灌木样方共两个,并随机设置两个1 m×1 m的草本样方;每个10 m×10 m样方内随机设置5 m×5 m的灌木样方和1 m×1 m的草本样方各一个,共有乔木样方36,灌木样方35,草本样方36个。乔木样方内对高度≥5 m的乔木进行每木调查,记录其种名、高度、胸径、枝下高、冠幅;灌木记录其种名、各物种株数、平均高度、平均冠幅、乔灌幼苗数;草本记录其种类、平均高度、平均盖度。采用GPS和地质罗盘测量各样地海拔、经纬度、坡度和坡向等。本次调查共记录到维管束植物288种,隶属于94科213属,其中:乔木82种,灌木197种,草本77种。
1.4 数据处理
计算各样方内每个物种的重要值,多样性的测度采用丰富度指数Margalef、多样性指数Shannon-Wiener、均匀度指数Pielou和优势度指数Simpson[19-20],计算公式为:
乔木重要值=(相对密度+相对频度+
相对显著度)/3
灌木重要值=(相对密度+相对高度+
相对盖度)/3
草本重要值=(相对高度+相对盖度)/2
丰富度Margalef指数:R=(S-1)/ln(N)
多样性Shannon-Wiener指数:H=-∑PilnPi
均匀度Pielou指数:J=H/lnS
式中:S——样地中记录到的物种数;N——样地内所有物种的重要值之和;Pi——物种i的重要值占样方全部物种重要值的比例。
利用Excel计算物种多样性指数,SPSS软件进行方差分析,Canoco 4.5软件对三种生长型植物的重要值(乔木82种、灌木197种、草本77种)与4个环境因子(海拔、坡度、坡向及坡位)分别整理成数据矩阵进行排序。坡向和坡位采用数字等级赋值表示,1表示阴坡,2表示半阴坡,3表示半阳坡,4表示阳坡;1表示山顶、2表示山脊、3表示上坡、4表示中坡、5表示下坡、6表示河谷,其他环境数据采用实测值表示。
2 结果与分析
2.1 地形对不同生长型植物物种多样性的影响
对于乔木(表1),不同坡位的多样性和丰富度指数无显著差异;山顶的均匀度指数最低且显著(p<0.05)低于其他坡位;优势度与均匀度指数相反,山顶最高且显著(p<0.05)高于其他坡位。坡向对四个多样性指数无显著影响。坡度在平坡处的多样性指数显著(p<0.05)高于缓坡和斜坡处,陡坡处的多样性显著(p<0.05)高于斜坡;丰富度指数与多样性指数的变化一致;均匀度指数为斜坡处显著(p<0.05)低于平坡和陡坡处;优势度指数为斜坡处显著(p<0.05)高于平坡和陡坡处。对于灌木(表2),在不同坡位,山顶的多样性指数要显著(p<0.05)高于下坡和河谷;丰富度指数为山顶显著(p<0.05)高于中坡、下坡和河谷,上坡显著(p<0.05)高于下坡和河谷;均匀度指数无显著差异;优势度指数为河谷显著(p<0.05)高于山顶。坡向和坡度对灌木的四个多样性指数无显著影响。对于草本(表3),山顶的多样性指数显著(p<0.05)低于中坡、下坡和河谷;丰富度指数为山顶和山脊显著(p<0.05)低于中坡、下坡和河谷;均匀度指数无显著差异;优势度指数为山顶显著(p<0.05)高于中坡、下坡和河谷。半阴坡的丰富度指数显著(p<0.05)高于半阳坡,其他指数无显著差异;坡度对草本的4个多样性指数无显著影响。
2.2 地形对不同生长型植物分布格局的影响
对乔木、灌木和草本植物与环境因子分别进行CCA排序,蒙特卡罗检验结果显示所有典范轴的显著性均达到显著水平(p<0.01)。在乔木、灌木和草本的CCA排序轴中,前3轴的物种环境方差累积贡献率分别是87.1%,81.7%,82.8%(表4)。前3轴的累积贡献率达80%,说明排序结果能够较好地解释物种与环境间的相关关系。
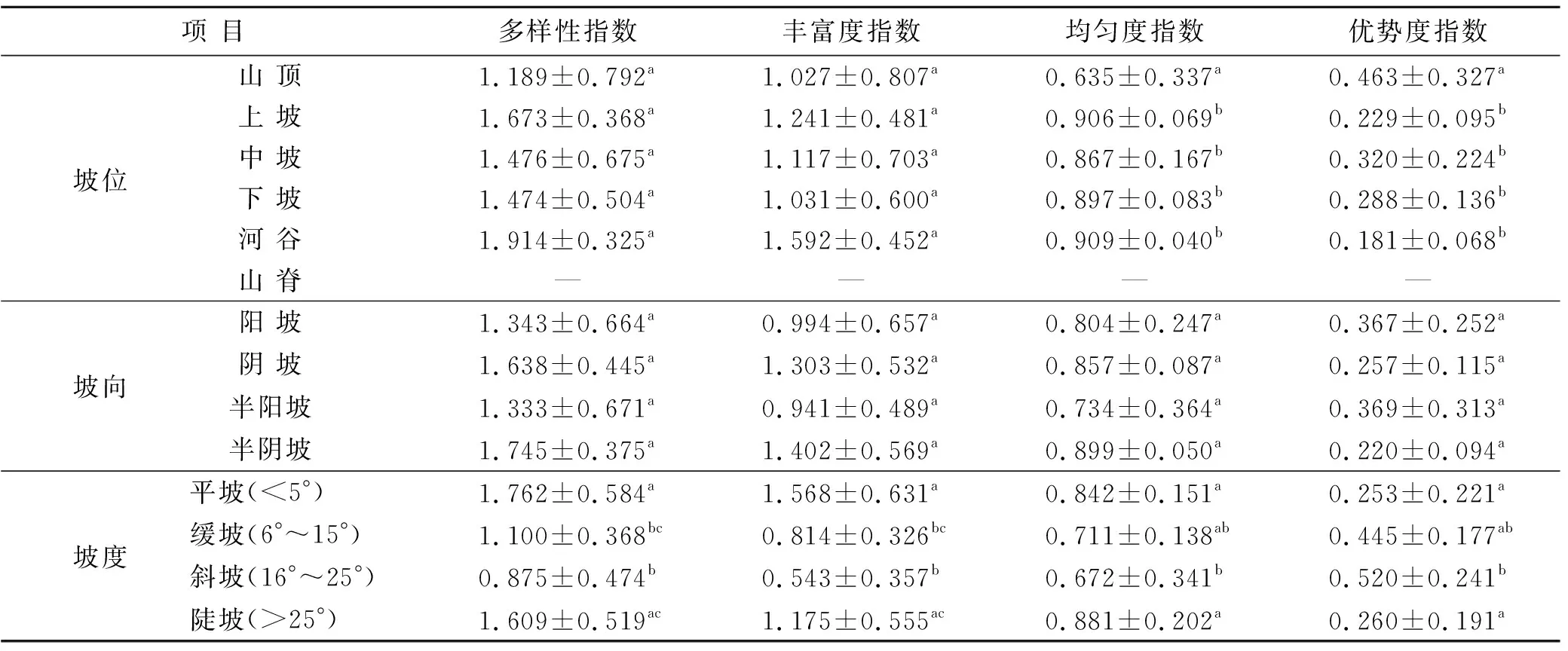
表1 不同地形条件下乔木物种多样性指数
注:“—”表示无数据; 表中数据为均值±标准误; 同列数据后不同字母表示差异显著(p<0.05)。下同。

表2 不同地形条件下灌木物种多样性指数

表3 不同地形条件下草本物种多样性指数

表4 3种生长型植物排序的特征值和物种-环境相关系数
注:CCA1,CCA2,CCA3分别表示第一排序轴、第二排序轴、第三排序轴。
由于前两个排序轴反映了物种与环境关系的绝大部分信息,所以利用前两轴绘制乔木、灌木和草本的物种—环境因子排序图。箭头表示环境因子,箭头的长短代表该环境因子与群落分布或物种分布的相关程度;箭头与排序轴夹角的大小表示该环境因子与排序轴的相关性[21]。不同物种对环境的要求存在差异,在排序图上的分布情况也会有所不同。在乔木排序图中(图1),4个地形因子中与第一排序轴相关性比较高主要有坡位和海拔,从左到右表示坡位由山顶、上坡、中坡、下坡到河谷,海拔越来越低。位于第一排序轴左端的物种主要分布在山顶、上坡和海拔较高的区域,包括马尾松(Pinusmassoniana)、大明松(Pinustaiwanensis)、乌冈栎(Quercusphillyraeoides)、檵木(Loropetalumchinense)、柏树(Cupressusfunebris)等;右端的物种多分布在中下坡和河谷等海拔较低的区域,主要包括小果润楠(Machilusmicrocarpa)、日本杜英(Elaeocarpusjaponicus)、红果黄肉楠(Actinodaphnecupularis(Hemsl.)Gamble)、朴树(Celtissinensis)、黄丹木姜子(Litseaelongata)等。与第二排序轴相关性较高的主要是海拔和坡向,从上到下海拔越来越低,坡向由阳坡、半阳坡、半阴坡到阴坡。在第二排序轴上方的物种多分布在海拔较高,坡位向阳的区域,主要有黄栌(Cotinuscoggygria)、岩生鹅耳枥(Carpinusrupestris)、海桐(Pittosporumtobira)、圆果化香(Platycaryalongipes)、杜鹃(Rhododendronsimsii)等;下方的物种多分布在低海拔、阴坡和半阴坡等区域,主要有冬青(Ilexchinensis)、珊瑚朴(Celtisjulianae)、榆树(UlmuspumilaL.)、青冈(Cyclobalanopsisglauca)、山胡椒(Linderaglauca)等。
在灌木排序图中(图2),由于物种种类较多,故图中只显示权重>2%的物种。图中与第一排序轴相关性比较高的地形因子主要有海拔和坡位,从左到右表示海拔越来越低,坡位由山顶、山脊、上坡、中坡、下坡到河谷。左端的物种主要位于山顶、山脊、上坡和海拔较高的区域,包括化香树(Platycaryastrobilacea)、铁仔(MyrsineafricanaL.)、绣线菊(SpiraeasalicifoliaL.)、火棘(Pyracanthafortuneana)、荚蒾(Viburnumdilatatum)等;右端的物种主要分布在低海拔和下坡、河谷等区域,主要有水麻(Debregeasiaedulis)、花椒簕(Zanthoxylumscandens)、苎麻(Boehmerianivea)、高粱泡(Rubuslambertianus)、窄叶蚊母树(Distyliumdunnianum)等。与第二排序轴相关性较高的主要是坡度,从上到下坡度越来越小。坡度较陡的区域主要分布有异叶鼠李(Rhamnusheterophylla)、砚壳花椒(Zanthoxylumdissitum)、小果蔷薇(Rosacymosa)、黄脉莓(Rubusxanthonearus)等;在坡度较为平缓的区域主要分布有悬钩子(Rubuscorchorifolius)、异叶梁王茶(Nothopanaxdavidii)、构树(Broussonetiapyrifera)、盐肤木(Rhuschinensis)等。

图1 乔木物种与环境因子的CCA排序图
在草本植物的排序图中(图3),与第一排序轴相关性较高的环境因子主要是海拔,其次是坡位,从左到右表示海拔越来越低,坡位由山顶、山脊、上坡、中坡、下坡到河谷。在山顶、山脊、上坡等海拔较高的区域主要分布有狗尾草(Setariauiridis)、苔草(Carex)、沿阶草(Ophiopogonbodinieri)、淫羊藿(Epimediumsagittatum)等;卷柏(Selaginellatamariscina)、新月蕨(Pronephriumgymnopteridifrons)、蒜香藤(Mansoaalliacea)等主要分布在下坡、河谷等海拔较低的区域。与第二排序轴相关性较高的主要坡度,从上到下坡度越来越小。在坡度较大的区域主要分布有贯众(Cyrtomiumfortunei)、蛇根草(OphiorrhizamungosL.)、冷水花(Pileanotata)等;在坡度较小的区域主要分布有金星蕨(Parathelypterisglanduligera)、鸢尾(Iristectorum)、香附子(Cyperusrotundus)等。
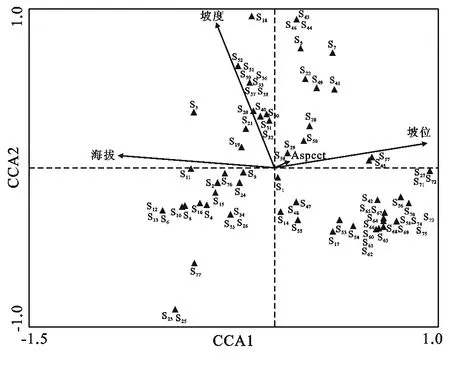
图3 草本物种与环境因子的CCA排序图
同时通过前向选择法分析各项环境因子对三种生长型植物的影响力和显著性(表5),由表可知,地形因子中对乔木分布格局的解释力度大小为:坡位>海拔>坡度>坡向,前三者影响比较显著;对于灌木,地形因子影响力排序为:海拔>坡度>坡位>坡向,前两者比较显著;对于草本,影响力排序为:海拔>坡度>坡向>坡位,具有显著影响力的是海拔和坡度。

表5 环境因子对植物分布的解释量与显著性检验
注:*表示在0.05水平上显著,**表示在0.01水平上显著。
3 讨 论
3.1 地形因子对植物物种多样性的影响
地形差异形成的异质生境是物种多样性维持的重要基础[3]。不同坡位土壤水肥条件具有显著差异,最终影响植物多样性的差异[22]。本研究中,不同坡位乔木的丰富度和多样性指数无显著差异;山顶由于光照充足,土壤温度较高,马尾松、大明松等阳性树种分布较多,优势种明显,优势度指数明显高于其他坡位,但均匀度指数明显低于其他坡位。在山顶和上坡区域,灌木的丰富度和多样性指数都比较高,而下坡和河谷地区较低。下坡等区域乔木分布密度大、群落郁闭度高,灌木生长所需的光照条件受到限制,因而物种丰富度和多样性指数显著低于其他坡位;河谷地段海拔较低,气候湿热,水流冲刷严重、石砾含量高且土壤多为砂土,植被主要以窄叶蚊母树为主,物种单一且分布较多,所以灌木的丰富度和多样性指数低,但优势度指数较高。山顶和山脊光照充足,土壤养分含量少,植被多为耐瘠喜阳物种,而中下坡和河谷区域光照有限,但生境水分含量高,植被多为耐荫喜湿物种。草本植物在中下坡及河谷的丰富度和多样性指数明显高于山脊和山顶,表明这一区域草本多为喜湿耐荫物种,少量喜阳物种多分布在山顶、山脊等区域,占较大的优势地位。
坡向影响地表植被接受太阳光照的时间和强度,导致土壤产生水热差异。彭晚霞等[23]认为坡向是导致土壤全氮和有机质含量产生变化的主要因素,对群落物种丰富度具有显著影响。本研究中,坡向对乔木和灌木的4个多样性指数均无显著影响,说明乔木和灌木对坡向的变化并不敏感。但草本的丰富度指数表现为半阴坡显著高于半阳坡,主要由于该区域耐荫性草本植物种类较多,此外半阴坡的水分和光照条件都比较适中;半阳坡地区岩石裸露的微生境较多,土壤水分养分易流失,对草本类近地表植物的丰富度影响比较大。
坡度对灌木和草本4个多样性指数均无显著影响,但在平坡和陡坡处,乔木的多样性指数、丰富度指数及均匀度指数都比较高,这与高策[9]等研究表明的坡度增加物种数减少的趋势并不一致。平坡地形平坦,有利于土壤养分、水分物质的积累,利于植被生长;陡坡地带地势险峻,人为干扰小,原生植被保存较好,物种丰富度和多样性都较高。缓坡斜坡地带多靠近旅游栈道、人行步道,人为干扰较大,可能对乔木的物种多样性有一定影响。
3.2 地形因子对不同生长型植物分布格局的影响
在区域和景观尺度上,地形是影响植被物种组成和分布格局的主要环境因子。地形通过对光照、水分、温度、湿度等环境因子的控制影响土壤肥力和水热资源的重新分配,从而影响植被的物种组成和分布状况[23]。有研究[24-25]认为,在区域尺度内,海拔是影响植物分布格局的主要因子;也有研究[26]表明坡向对植物分布格局的影响大于海拔;而黄甫昭等[27]对弄冈喀斯特山地的研究表明海拔和坡向的变化是影响植被差异的共同因素。此外,有研究表明,在一定海拔范围内,坡位是影响植被生长的关键因子[28];而袁铁象等[29]在桂西南的研究表明坡度是影响灌木、草本等地表植物的分布格局的主要地形因子,其次是海拔和坡向。由于研究对象和取样尺度的差异,影响植被分布格局的地形因子往往不同[30]。
本研究中,对乔木物种分布格局具有显著影响的地形因子依次是坡位、海拔和坡度;海拔和坡度对灌木和草本物种的分布格局均具有显著影响,这与熊斌梅等[31]的研究结果相似。不同生长型植物对地形因子变化的响应有所差异,灌木和草本对坡位变化引起的生境改变的响应没有乔木敏感;云台山地势陡峭,山峰高耸,海拔是影响其水热条件差异的主要环境因子,植被分布呈明显的地形分异现象[16];以物理崩塌为主的地貌发育导致这一区域山体十分陡立,坡度对土壤水分、养分的作用加强,成为影响植物分布的主要因子之一;相比之下,坡向对几种生长型植被分布格局的影响并不显著。
4 结 论
坡位对乔木、灌木和草本的物种多样性均存在一定程度的影响。坡向对乔木、灌木的4个多样性指数影响不显著;草本在半阴坡的丰富度明显高于半阳坡。乔木在平坡和陡坡处的多样性指数和丰富度指数最大,平坡和陡坡处均匀度指数显著高于斜坡,优势度指数与均匀度指数相反;坡度对灌木和草本的4个物种多样性指数无显著影响。不同生长型植物对地形因子变化的响应不同,坡位、海拔和坡度是影响乔木物种分布格局的主要因子;而海拔和坡度是影响灌木和草本物种分布格局的主要因子,坡向对3种生长型植物分布格局的影响均不显著。云台山喀斯特区植物物种多样性及其分布格局受到以海拔、坡度为主,坡位和坡向为辅的多个因子共同的影响。
