南方红壤丘陵区土壤细菌对土壤水分和温度的响应差异
2019-06-11杨杰陈闻侯海军陈春兰秦红灵吕殿青
杨杰 陈闻 侯海军 陈春兰 秦红灵 吕殿青
摘 要 红壤丘陵区稻田不同时期进行的水分管理导致土壤水分和温度同时变化,继而影响土壤微生物群落结构,二者对土壤微生物的影响机制目前还不清楚。本研究以中国科学院桃源农业生态实验站长期定位试验的稻田作为研究对象,同时以旱地农田作为对照,利用末端限制性酶切片段长度多态性分析(T-RFLP)和荧光定量PCR(qPCR)方法,研究不同水分管理时期细菌群落结构与丰度的变化特征,进而分析其变化的影响因素,阐明水分和温度的相对贡献。结果表明,稻田土壤细菌受土壤水分状况的显著影响,淹水期土壤细菌群落结构明显区别于其他时期,且淹水期土壤细菌丰度和多样性指数均显著低于晒田期。稻田土壤细菌群落结构和多样性指数均受到土壤含水量和土壤温度的显著影响,但土壤含水量的相关性大于土壤温度。而作为对照的旱地农田土壤含水量在不同时期没有差异,土壤细菌群落结构也没有差异,仅土壤细菌丰度和多样性指数发生一定变化,但这种变化并不与土壤温度呈现相关性。因此,本研究认为,南方丘陵区农田土壤细菌对土壤水分的响应比对土壤温度更敏感。
关键词 稻田;旱地;细菌群落结构;细菌丰度;土壤水分;土壤温度
中图分类号 X171.3 文献标识码 A
红壤是我国重要的土壤资源,主要分布于长江以南的低山丘陵区,涉及诸多省区,其农业产值负担了全国近一半的人口[1],是中国粮食和亚热带经济作物及果、林、草发展的重要基地[2]。但是南方丘陵区雨水较多,是中国水土流失最为严重的区域之一[1],从而导致水土退化范围广、程度高,生态环境遭到严重破坏[3]。土壤微生物是生态系统的重要组成部分,是土壤碳、氮、磷等元素生物地球化学循环的主要推动力,在维持土壤生态系统的稳定性与功能方面起着关键作用[4-6]。
稻田是亚热带红壤区最主要的农田利用方式,当地农民广泛应用间歇灌溉的水分管理方式来增加水稻有效分蘖和水稻植株的光合效率,提高根系活力和根系对土壤养分的利用效率[7]。不同水分管理方式通过改变土壤相关物理化学性质,包括土壤水分、通气状况、Eh等,进而影响微生物种群[8-9]。例如张敬智等[10]研究发现,淹水稻田土壤微生物群落结构显著不同于好气条件下的稻田土壤。在农田生态系统中,土壤微生物的群落结构与数量可以作为衡量土壤肥力高低和土壤可持续生产力的重要指标。目前大部分的研究主要是依托野外定位试验或室内模拟试验,采集相同时间点不同水分处理土壤研究水分管理对土壤微生物群落结构的影响[11-12]。但在实际农业生产中,稻田间歇灌溉发生在不同时期,稻田土壤水分和温度同步发生变化。而土壤温度也是影响微生物群落结构的重要因素[13-14]。目前,不同水分管理措施下水分和温度对土壤微生物变化的影响机制还不清楚。而与稻田相比,亚热带红壤区旱地农田往往无人工灌溉,土壤水分含量保持稳定,而土壤溫度随季节而变化,为研究土壤微生物对水分和温度的响应方式提供了良好的土壤对照条件。
本研究采用T-RFLP和qPCR技术相结合的方法,以红壤丘陵区近30年一直种植双季稻的稻田土壤为对象,研究了不同采样时期土壤细菌丰度和群落结构的变化特征及其影响因素,同时以开垦年代相似、玉米-油菜轮作的旱作农田为对照,测定同时期的细菌丰度和群落结构特征。对比分析了土壤细菌群落对土壤水分和温度的敏感性,以期阐明不同水分管理措施下红壤稻田土壤细菌丰度和群落结构变化的驱动机制,为认识不同管理条件下稻田土壤微生物的特征及制定农田合理的水分管理措施等提供基础数据。
1 材料与方法
1.1 材料
1.1.1 供试土壤 土壤样品采自中国科学院桃源农业生态试验站。该试验场地处湖南省桃源县漳江镇宝洞峪村 (111°26'E,28°55'N),气候属于中亚热带向北亚热带过渡的季风湿润气候,年均气温16.5 ℃,年均降水量为1447.9 mm,年日照时数1530 h,土壤类型为第四纪红土黏土发育的红壤,稻田以种植双季稻为主,旱地主要种植玉米、油菜等。
1.1.2 仪器与设备 流动注射仪,型号AA3,德国SEAL公司;有机碳分析仪,型号Vwp,日本岛津公司;荧光定量PCR仪,型号ABI 7900 HT,美国AB公司。
1.2 方法
1.2.1 试验设计 在桃源农业生态试验站长期定位试验田中,选择双季稻和玉米-油菜轮作旱地两种农田利用方式,分别设置3个独立小区(8 m×5 m)作为重复样地。双季稻田全年施肥量为:N 182.3 kg/hm2, P 39.3 kg/hm2, K 197.2 kg/hm2;玉米-油菜轮作旱地施肥量为:N 240 kg/hm2, P 52.4 kg/hm2, K 286.4 kg/hm2。双季稻田在水稻生育期进行淹水-烤田-淹水-干湿交替的水分管理模式,淹水保存田间水层5 cm,晒田排水后自然晒干2周;玉米-油菜轮作旱地没有人工灌水。
1.2.2 样品采集 2012年10月—2014年7月,在稻田淹水期(7月,夏季;F)、晒田期(10月,秋季;D)和冬春干旱期(1月和4月;W,S)采集稻田土壤样品,同时间采集同时期旱地土壤样品作为对照,不同年际样品作为采样重复。为了尽量减少施肥对土壤性状的影响,样品采集于施肥前或施肥2月后。2014年7月,稻田采样时,旱地刚刚施肥一周,因此,本研究剔除了这次的旱地样品。每个小区S形多点采集表层0~ 20 cm土壤后混匀,一部分土壤(100 g)迅速用液氮冰冻后80 ℃保存供分子生物学研究,另一部分土样(400 g)4 ℃保存用于测定土壤铵态氮、硝态氮和土壤可溶性有机碳(DOC)等相关理化性质。2012年采样开始时,土壤基本理化性状见表1。
1.2.3 数据收集 土壤理化性质测定:采样当天,利用地温计测定5 cm土层土壤温度;土壤含水量使用烘干法;土壤铵态氮和硝态氮用1 mol/L的KCl溶液浸出的土壤悬浮液,过滤后使用流动注射系统进行分析;土壤可溶性有机碳(DOC)利用0.5 mol/L的K2SO4浸提,过滤后用有机碳分析仪测定。稻田土壤硝态氮含量低于检测线,不进行后续分析。
微生物丰度及群落结构的测定:DNA的提取参照杨建等[15]的SDS-GITCPEG方法。荧光定量PCR采用采用16S通用引物1369F/1492R在荧光定量PCR 仪上进行自动化PCR 反应定量扩增细菌16S rDNA,引物信息及PCR反应条件同秦红灵等[4]。标准曲线的建立参照Chen等[16]的方法。T-RFLP试验采用16S通用引物27F/1492R扩增细菌16S rDNA,其中上游引物5'端用6-羧基二乙酸荧光素(FAM)标记,用于T-RFLP试验[4]。末端荧光标记的PCR纯化后产物用 MspⅠ消化,酶切产物的T-RFLP分析由上海桑尼生物技术有限公司完成。
1.3 数据处理
所有测定结果均为3次重复的平均值,误差线为标准差(SD)。使用SPSS 10.0软件单因素方差分析(One-Way ANOVA)和多重比较分析(LSD)不同采样时期土壤理化性质以及细菌丰度和多样性的差异,Pearson相关分析土壤理化性质与土壤细菌丰度和多样性指数的关系。利用Canoco v. 4.5软件主成分分析(PCA)不同采样时期土壤细菌群落结构的相似性,蒙特卡洛检验土壤理化性质对细菌群落结构的影响。
2 结果与分析
2.1 红壤稻田和旱地土壤理化性状
方差分析结果表明,稻田土壤含水量、土壤温度、铵态氮和DOC含量均受采样时期的影响。稻田夏季淹水期土壤含水量、土壤温度和铵态氮含量显著高于其他3个时期,但DOC含量显著低于其他时期;而旱地无人为灌溉,不同采样时期土壤含水量没有差异,但土壤温度呈现了和稻田相同的变化规律,另外土壤硝态氮、铵态氮和DOC含量不同采样时期也存在显著差异,冬季土壤铵态氮和土壤DOC最高,秋季土壤硝态氮含量最高(表2)。
2.2 红壤稻田和旱地土壤细菌群落结构
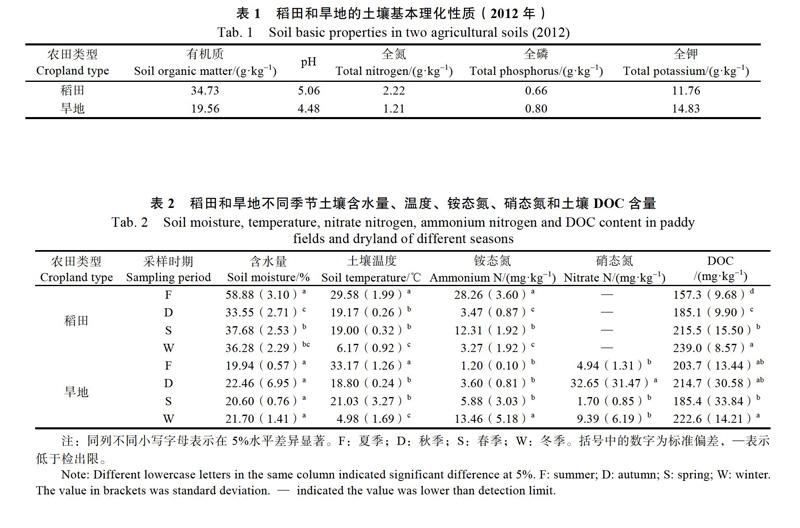
根据T-RFLP 图谱分析发现,稻田土壤细菌群落结构受采样时期的显著影响,夏季淹水期土壤细菌分布明显与其他3个时期不同。淹水期相对丰度排在前3位的bp片段分别是:150、148和436 bp t-RFs,其中,150 bp t-RF在淹水期相对丰度最高为30.79%,比其他3个采样时期高出10%以上;436 bp t-RF淹水期相对丰度为9.53%,是其他3个时期的2倍左右(图1A)。利用网络在线服务(http://trflp.limnology.wisc.edu/index. jsp)PAT工具(The T-RFLP Phylogenetic Assignment Tool)可以得到,150 bp與甲烷氧化菌(AF150788)末端片段一致,436 bp与海洋雪相关细菌(AF030780)末端片段一致。主成分分析(PCA)也表明,稻田淹水期土壤细菌在PC1轴与其他3个采样时期明显分离(图1C)。而旱地土壤细菌群落结构不受采样时期的影响,土壤细菌群落交错分布,没有明显的聚类(图1B,图1D)。
2.3 红壤稻田和旱地土壤细菌丰度和多样性指数
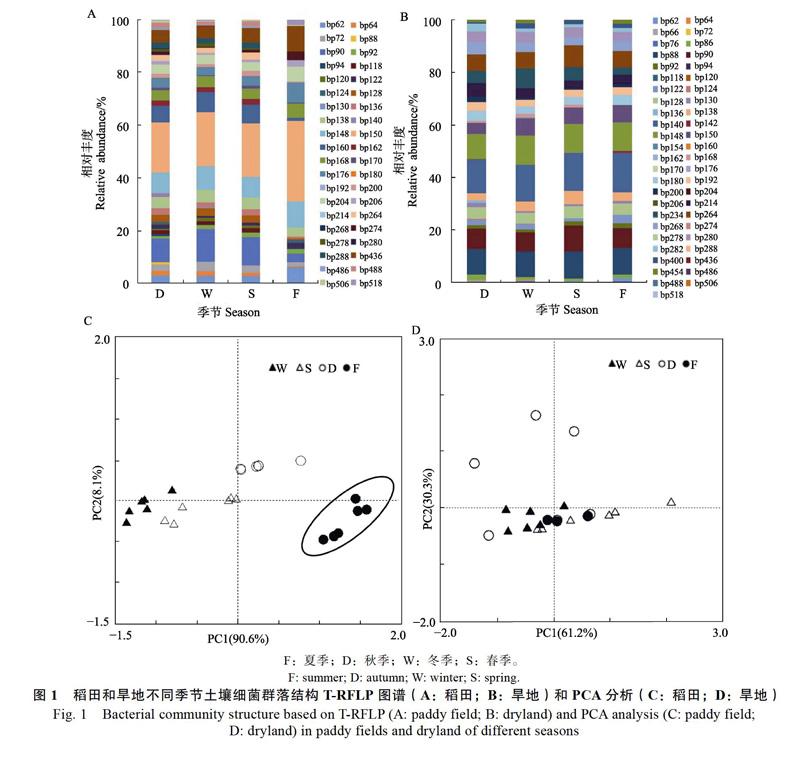
方差分析结果表明,稻田土壤细菌丰度和多样性指数均受到采样时期的影响,夏季淹水期土壤丰度和多样性指数低于秋季晒田期,冬春旱季干旱期居中(图2A,图2C)。旱地土壤细菌丰度和多样性指数也受到采样时期的影响,但两者的趋势不一致,秋季土壤细菌丰度最低,多样性指数春季较低(图2B,图2D)。
2.4 土壤细菌群落结构、丰度及多样性与土壤理化性质的关系
蒙特卡洛检验发现,稻田土壤细菌群落结构受土壤含水量、土壤温度、铵态氮和DOC含量的
影响,而旱地土壤群落结构只受到土壤DOC含量的影响;与群落结构结果相一致,稻田土壤细菌多样性指数受到土壤含水量、土壤温度和铵态氮的影响,但细菌丰度与测定的土壤理化性质没有显著相关关系;旱地土壤细菌丰度和多样性指数与测定的土壤理化性质均不存在显著相关关系(表3)。
3 讨论
3.1 土壤细菌群落结构对土壤水分的响应比温度更敏感
间歇灌溉是亚热带红壤区稻田水分管理的常见方式,而稻田水分管理方式对土壤微生物具有显著的影响。本研究采集不同水分管理时期的稻田土壤样品,研究稻田不同水分状态对土壤细菌群落的影响发现,淹水期稻田土壤细菌群落结构明显区别于其他3个采样时期,且淹水期土壤细菌多样性指数和丰度均小于晒田期。稻田淹水不仅造成土壤含水量的降低,同时还意味着土壤Eh
和氧气浓度的降低,抑制了土壤中好气微生物的生长[17],同时促进了某些厌氧微生物的生长,如反硝化菌和产甲烷菌[18-19],从而造成了细菌群落组成和多样性的变化。同时,Sahrawat[20]和张薇等[21]研究发现,淹水条件不利于土壤有机碳的矿化,抑制了土壤的供碳能力,这一方面影响细菌的总体生长状况,另一方面也会影响微生物的组成。例如张敬智等[10]研究发现,淹水稻田和好气稻田由于有机碳组分的不同,微生物群落结构产生适应性变化。本研究中淹水期土壤细菌丰度和多样性的减少,可能与特定功能微生物的富集有
表3 土壤细菌丰度、多样性指数及群落结构与土壤理化性质的关系
Tab. 3 Relationships between soil bacterial abundance, diversity, community structure and soil physicochemical properties
指标
Items 农田类型
Cropland type 相关性
Correlation 土壤含水量 Soil moisture 土壤温度
Soil temperature 土壤铵态氮
Ammonium N 土壤硝态氮 Nitrate N DOC
细菌丰度 稻田 R 0.32 0.18 0.37 — 0.15
ρ 0.13 0.41 0.08 — 0.48
旱地 R 0.18 0.15 0.16 0.20 0.37
ρ 0.44 0.52 0.48 0.39 0.10
细菌多样性指数 稻田 R 0.58 0.44 0.61 — 0.26
ρ 0.00* 0.03* 0.00* — 0.22
旱地 R 0.25 0.18 0.20 0.34 0.22
ρ 0.28 0.44 0.39 0.13 0.35
細菌群落结构 稻田 R 14.12 10.13 14.69 — 6.18
ρ 0.00* 0.00* 0.00* — 0.01*
旱地 R 0.88 0.69 0.60 1.54 3.43
ρ 0.42 0.65 0.68 0.14 0.02*
注:细菌丰度和多样性与土壤性质的关系采用Pearson相关分析,细菌群落结构与土壤性质的关系采用蒙特卡洛检验;样本数:稻田n=24;旱地n=21。*表示相关性达到5%的显著水平。
Note: Pearson correlation analysis was used to analyze the relationship between soil bacterial abundance and soil properties, and Monte Carlo method was used to analyze the relationship between community structure and soil properties. Number of samples: paddy field n=24; dryland n=21. * indicated the correlation reached 5% significance level.
关,T-RFLP图谱分析也确实发现了某些与土壤水分含量相关的功能微生物如甲烷氧化菌和海洋雪相关细菌在稻田富集。
温度也是造成土壤微生物改变的重要因素,温度和土壤微生物的关系已被很多研究者所证明[13, 22]。例如Fey等[23]通过室内设置不同温度梯度,研究发现温度对水稻土产甲烷古菌的群落结构、活性和功能均产生显著影响。但是在本研究中,土壤温度对土壤微生物的影响较小,例如稻田冬春干旱期,土壤含水量没有差异,土壤温度相差12.83 ℃,但是土壤细菌群落结构、丰度和多样性指数均没有显著差异;旱地夏季和冬季采样时期土壤水分含量没有差异,温度相差28.19 ℃,土壤细菌群落结构、丰度和多样性指数也没有显著差异。土壤水分对微生物的影响强度高于温度,在其他研究中也有发现,例如亚热带森林土壤海拔高度改变时,微生物群落功能多样性产生差异的主要原因是土壤含水量,而非土壤温度[24]。另外,土壤温度与土壤微生物的相关性可能需要基于良好的水分环境,例如李世清等[25]的研究发现,当田间土壤含水量足够维护微生物的活性,土壤温度与土壤微生物量氮之间存在显著的正相关关系。而在本研究中,旱地农田土壤水分含量稳定,且显著低于稻田水分含量,因此土壤温度与土壤微生物并没有表现出显著的相关性,而稻田中二者却表现出了正相关关系。因此,本研究证实,在亚热带红壤区,土壤水分而非土壤温度是调控农田生态系统微生物群落结构的关键因子。
3.2 稻田与旱地土壤群落结构的差异原因分析
稻田与旱地由于种植方式的不同,土壤微生物群落结构产生显著差异。本研究发现旱地土壤中大于10%相对丰度的优势菌群(148、140 bp t-RFs)不同于水田土壤(150 bp t-RF)。稻田和旱地农田土壤微生物的不同也被其他研究者证实。沈冰洁等[5]研究发现旱地土壤中α-变形菌和γ-变形菌的相对丰度较高,而稻田土壤中β-变形细菌和甲烷菌的相对丰度较高。李忠佩等[26]的研究也表明中国亚热带丘陵地区,稻田土壤微生物群落功能多样性高于旱地土壤。亚热带红壤区稻田土壤由于淹水、晒田等水分管理措施的存在,水分状态频繁发生变化,并且诱发了土壤通气状态、养分状态等的改变,而土壤微生物对氧气状态[18]和矿质氮形态[27]等具有偏好性,因此造成了稻田和旱地农田土壤微生物的不同。例如稻田土壤水分含量更高,可能更有利于厌氧或兼性厌氧微生物的生存,例如反硝化微生物[18]的生存;而旱地土壤水分含量较低,可能更有利于好气微生物的生存[17];稻田土壤矿质氮以铵态氮为主,可能更有利于喜铵微生物的生存,旱地以硝态氮为主,可能更有利于对硝态氮具有偏好性的微生物的生存。另外,土壤中功能微生物也会因为稻田和旱地农田土壤性质的不同而发生变化,例如铵态氮和硝态氮作为硝化作用和反硝化作用的底物,稻田和旱地土壤间的差异,势必会引起硝化功能和反硝化功能微生物的差异。最终在这些因素的共同作用下,稻田和旱地土壤微生物产生了显著分异。
4 结论
南方红壤区稻田生态系统土壤细菌群落结构、丰度和多样性随采样时期土壤水分状况的不同产生显著差异,淹水期土壤某些功能微生物的富集,减少了土壤细菌丰度和多样性。旱地不同采样时期土壤含水量没有差异,即使夏冬采样时期土壤温度相差超过25 ℃,土壤细菌群落结构、丰度和多样性均没有显著差异。因此,本研究认为亚热带红壤丘陵区农田土壤细菌对土壤水分的响应比对土壤温度更敏感。
参考文献
程冬兵. 南方红壤丘陵区水土流失综合治理技术[C]//中国水土保持学会, 台湾中华水土保持学会. 2014年海峡两岸水土保持学术研讨会论文集. 武汉: 中国水土保持学会, 台湾中华水土保持学会, 2014: 6.
张 慧, 袁红朝, 朱亦君, 等. 不同利用方式对红壤坡地微生物多样性和硝化势的影响[J]. 生态学杂志, 2011, 30(6): 1169-1176.
黄河仙, 谢小立, 王凯荣, 等. 不同覆被下红壤坡地地表径流及其养分流失特征[J]. 生态环境, 2008, 17(4): 1645-1649.
秦红灵, 袁红朝, 张 慧, 等. 红壤坡地利用方式对土壤细菌群落结构的影响[J].土壤学报, 2011, 48(3): 594-602.
沈冰洁, 祝贞科, 袁红朝, 等. 不同种植方式对亚热带红壤微生物多樣性的影响[J]. 环境科学, 2015, 36(10): 3839-3844.
黄梦青. 土地利用方式及施氮对中亚热带山地土壤微生物生物量和群落结构的影响[D]. 福州: 福建师范大学, 2013.
林贤青, 周伟军, 朱德峰, 等. 稻田水分管理方式对水稻光合速率和水分利用效率的影响[J]. 中国水稻科学, 2004, 18(4): 55-60.
Pan F X, Li Y Y, Chapman S J, et al. Effect of rice straw application on microbial community and activity in paddy soil under different water status[J]. Environmental Science and Pollution Research, 2016, 23(6): 5941-5948.
侯海军, 张文钊, 沈建林, 等. 水分管理对稻田细菌丰度与群落结构的影[J]. 生态环境学报, 2016, 25(9): 1431-1438.
张敬智, 马 超, 郜红建. 淹水和好气条件下东北稻田黑土有机碳矿化和微生物群落演变规律. 农业环境科学学报, 2017, 36(6): 1160-1166.
Liu R, Hayden H L, Suter H, et al. The effect of temperature and moisture on the source of N2O and contributions from ammonia oxidizers in an agricultural soil[J] . Biology and Fertility of Soils, 2017, 53(1): 141-152.
Qin H, Tang Y, Shen J, et al. Abundance of transcripts of functional gene reflects the inverse relationship between CH4 and N2O emissions during mid-season drainage in acidic paddy soil[J]. Biology and Fertility of Soil, 2018, 54(8): 885–895
杨 林, 陈亚梅, 和润莲, 等. 高山森林土壤微生物群落结构和功能对模拟增温的响应[J]. 应用生态学报, 2016, 27(9): 2855-2863.
王学娟, 周玉梅, 江肖洁, 等. 增温对长白山苔原土壤微生物群落结构的影响[J]. 生态学报, 2014, 34(20): 5706-5713.
杨 建, 洪 葵. 红树林土壤总DNA不同提取方法比较研究[J]. 生物技术通报, 2006(S1): 366-371.
Chen Z, Liu J, Wu M, et al. Differentiated response of denitrifying communities to fertilization regime in paddy soil[J]. Microbial Ecology, 2012, 63(2): 446-459.
刘艳丽, 张 斌, 胡 锋, 等. 干湿交替对水稻土碳氮矿化的影响[J]. 土壤, 2008, 40(4): 554-560.
Liu J, Hou H, Sheng R, et al. Denitrifying communities differentially respond to flooding drying cycles in paddy soils[J]. Applied Soil Ecology, 2012, 62: 155-162.
吴 讷, 侯海军, 汤亚芳, 等. 稻田水分管理和秸秆还田对甲烷排放的微生物影响[J]. 农业工程学报, 2016, 32(s2): 69-76.
Sahrawat K L. Organic matter accumulation in submerged soils[J]. Advances in Agronomy, 2003, 81(3): 169-201.
张 薇, 王子芳, 王 辉, 等. 土壤水分和植物残体对紫色水稻土壤有机碳矿化的影响[J]. 植物营养与肥料学报, 2007, 13(6): 1013-1019.
Allison S D, Treseder K K. Warming and drying suppress microbial activity and carbon cycling in boreal forest soils[J]. Global Change Biology, 2008, 14(12): 2898-2909.
Fey A, Conrad R. Effect of Temperature on carbon and electron flow and on the archaeal community in methanogenic rice field soil[J]. Applied & Environmental Microbiology, 2000, 66(11): 4790-4797.
宋贤冲, 郭丽梅, 田红灯, 等. 猫儿山不同海拔植被带土壤微生物群功能多样性[J]. 生态学报, 2017, 37(16): 5428-5435.
李世清, 任书杰, 李生秀. 土壤微生物体氮的季节性变化及其与土壤水分和温度的关系[J].植物营养与肥料学报, 2004, 10 (1): 18-23.
李忠佩, 吴晓晨, 陈碧云. 不同利用方式下土壤有机碳转化及微生物群落功能多样性变化[J]. 中国农业科学, 2007, 40(8): 1712-1721.
陈晓光, 李洪民, 张爱君, 等. 氮素形态对甘薯土壤微生物及酶活性的影响[J]. 西南农业学报, 2017, 30(3): 619-623.
