不同种源枫香树幼苗生长性状的地理变异
2019-06-11何庆海李文鑫谢宇凯张燕琴石从广杨少宗
何庆海, 方 茹, 李文鑫, 谢宇凯, 张燕琴, 石从广, 杨少宗,①
(1. 浙江省林业科学研究院, 浙江 杭州 310023; 2. 中南林业科技大学, 湖南 长沙 410004)
种源实验是植物遗传育种的重要手段,用于研究种群遗传变异与环境因子的关系,阐明种群的变异模式[1]。 种源试验已被广泛应用于林木育种研究,在花楸树〔Sorbuspohuashanensis(Hance) Hedl.〕[2]、浙江楠(PhoebechekiangensisC. B. Shang)[3]和暴马丁 香〔Syringareticulatavar.amurenis( Rupr.)Pringle〕[4]等种类的研究中已取得一定成果。 由于林木遗传育种周期较长,苗期试验成为林木优良种源初选的重要方法, 并已在麻栎(QuercusacutissimaCarruth.)[5]、欧洲花楸(SorbusaucupariaLinn.)[6]和红椿(ToonaciliataRoem.)[7]等植物的研究中予以应用。
枫香树(LiquidambarformosanaHance)隶属于金缕梅科(Hamamelidaceae) 枫香树属(LiquidambarLinn.),是第三纪孑遗植物,在第四纪冰川于东亚避难所中得以保存[8]。 枫香树在中国主要分布于秦岭—淮河以南地区,跨北热带、南亚热带、中亚热带和北亚热带4 个气候带[9]。 在枫香树的广大分布区范围内,环境的异质性对不同种源枫香树产生的影响存在差异。 表型性状变异受遗传和环境因子二者的共同作用[10-11]。 Sun 等[12]认为,地理隔离并不是造成枫香树遗传差异的主要原因,枫香树的遗传多样性更多存在于种群内不同个体间。 已有研究通过种源试验观测1 年生、4.5 年生、10 年生和14 年生枫香树株高、胸径和积材等生长性状,并通过比较种源间的差异选择出优良种源和家系[13-16],从时空角度展示了不同生长年份枫香树种源间的差异。 而明确枫香树种源间生长性状差异的形成原因是选择优良种质资源的关键条件。
目前,地理-气候因子对枫香树种源的影响尚未明确,枫香树的苗期生长模型也亟需完善。 地理-气候因子作为可以测量的因子,在选择优良种质上具有重要的参考价值。 枫香树苗期生长模型的建立,不仅有利于了解其不同生长阶段的生长情况以及比较种源间的差异,还可为速生种质的选择提供有效的数据支撑。
本研究以30 个种源的枫香树种子为材料,通过播种繁殖获得实生苗,并分阶段测量1 年生幼苗的株高、地径和叶片形状因子,比较不同种源枫香树不同阶段的差异性,建立苗期生长模型;并结合种源地的地理-气候因子,分析影响不同种源枫香树生长的关键因子,探索其生长性状在地理空间上的变化规律,以期为选择适生的速生种质提供苗期数据,并为探索枫香树的地理变异奠定基础。
1 材料和方法
1.1 材料
于2014 年9 月至12 月,在15 个省(自治区),以县域范围较为集中的枫香树天然群体为种源,共收集30 个种源,每个种源枫香树单株均在10 株以上,株距在100 m 以上。 选择生长健壮且结实状况良好的单株,每个单株采集果实5 kg 以上,取出的种子于-20 ℃保存。
于2015 年4 月,在长乐林场(北纬30.37°、东经119.84°,海拔37 m)穴盘育苗,采用随机区组实验设计,重复3 次,每次100 粒种子,每个种源共300 粒种子。 常规水肥管理,出苗初期遮阳。 供试枫香树种源地的地理-气候因子见表1。
1.2 方法
每个重复随机选择15 株枫香树幼苗并标记,每个种源共45 株。 分别于2015 年的8 月和12 月以及2016 年3 月测量株高和地径,其中,使用皮尺(精度0.1 cm)测量株高,使用游标卡尺(精度0.001 mm)测量地径(距地面5 cm 处)。 于2015 年9 月下旬,每个重复随机选择15 株枫香树幼苗,每个种源共45株,取从上往下第3 或第4 枚叶,用YMJ-D 叶面积测量仪(浙江托普仪器有限公司)测量叶面积(a)和叶周长(p),并据此计算叶片形状因子(f),计算公式为f=4πa/(p2)。

表1 供试枫香树种源地的地理-气候因子Table 1 Geographical-climatic factors of provenance locations of Liquidambar formosana Hance tested
1.3 数据处理
根据最后1 次测量的数据计算各种源枫香树幼苗株高、地径和叶片形状因子的变异系数。 对种源间株高、地径和叶片形状因子进行方差分析,采用Duncan's新复极差法进行多重比较。 对生长性状与种源地的地理-气候因子进行相关性分析。 采用逐步回归法分析地理-气候因子对生长性状的影响,并建立线性回归方程[17]。 利用EXCEL 2010 和SPSS 19.0 软件进行数据处理与分析,利用Origin 8.5 软件绘图。
2 结果和分析
2.1 幼苗生长性状的比较
不同种源枫香树幼苗不同阶段株高和地径的比较结果见图1,不同种源枫香树幼苗叶片形状因子的比较结果见图2。

图1 不同种源枫香树幼苗不同阶段株高和地径的比较Fig. 1 Comparison on plant height and ground diameter at different stages of Liquidambar formosana Hance seedlings from different provenances
2.1.1 对株高和地径的影响 由图1 可见:第1 阶段(2015 年4 月至8 月),30 个枫香树种源幼苗株高的变化幅度较小,其中,贵州德江(P26)、陕西宁强(P28)和陕西镇巴(P30)种源的株高较大。 第2 阶段(2015 年8 月至12 月),30 个枫香树种源幼苗株高的变化幅度较大,其中,海南黎母山(P1)、海南霸王岭(P2)和云南富宁(P7)种源的株高较大。 第3 阶段(2015年12 月至2016 年3 月),30 个枫香树种源幼苗株高的变化幅度均较小。 2016 年3 月,30 个枫香树种源中P2 种源的株高最高,为65.0 cm,河南商城(P27)种源的株高最低,为25.5 cm。
与株高相比较,不同阶段枫香树幼苗地径在种源间的变化幅度均较小。 2016 年3 月,30 个种源中安徽霍山(P19)种源的地径最大,为6.478 mm,湖北京山(P23)和福建华安(P10)种源的地径较小,分别为4.635 和4.671 mm。
方差分析结果表明:枫香树幼苗株高和地径在种源间存在极显著差异。
2.1.2 对叶片形状因子的影响 由图2 可见:P7 种源枫香树幼苗叶片形状因子最大,为0.44,江苏苏州(P14)种源的叶片形状因子最小,为0.34。 方差分析结果表明:枫香树幼苗叶片形状因子在种源间存在显著差异。

图2 不同种源枫香树幼苗叶片形状因子的比较Fig. 2 Comparison on leaf shape factor of Liquidambar formosana Hance seedlings from different provenances
2.2 幼苗生长性状变异系数的比较
不同种源枫香树幼苗生长性状变异系数的比较结果见表2。 由表2 可见:30 个枫香树种源幼苗生长性状变异系数的均值由高到低依次为株高(23.01%)、 地 径( 19.35%)、 叶 片 形 状 因 子(17.51%)。 30 个枫香树种源幼苗株高的变异系数的变化幅度较大,其中,福建尤溪(P9)种源株高的变异系数最大(33.90%),海南霸王岭(P2)种源株高的变异系数最小(8.79%),前者约为后者的4 倍;30 个枫香树种源幼苗地径的变异系数的变化幅度较小,其中,陕西镇巴(P30) 种源地径的变异系数最大(24.56%),安徽霍山(P19)种源地径的变异系数最小(15.37%);30 个枫香树种源幼苗叶片形状因子的变异系数的变化幅度较大,其中,江西大余(P16)种源叶片形状因子的变异系数最大(28.01%),P2 种源叶片形状因子的变异系数最小(8.90%),前者约为后者的3 倍。 种源内枫香树幼苗生长性状变异系数均值最大的是湖南桑植(P20)种源(25.43%),最小的是P2 种源(11.86%),表明不同种源枫香树幼苗生长性状种源内的变异存在明显差异。
2.3 相关性分析
枫香树幼苗生长性状与其种源地地理-气候因子间的相关系数见表3。 由表3 可见:枫香树幼苗株高与纬度和经度呈极显著(p<0.01)负相关,与海拔、年均温、1 月均温和无霜期呈极显著正相关,与年降水量无显著相关性;地径与纬度呈极显著正相关,与年均温、1 月均温和无霜期呈极显著负相关,与经度、海拔和年降水量无显著相关性;叶片形状因子与纬度呈极显著负相关,与年均温、1 月均温和无霜期呈极显著正相关,与经度、海拔和年降水量无显著相关性。
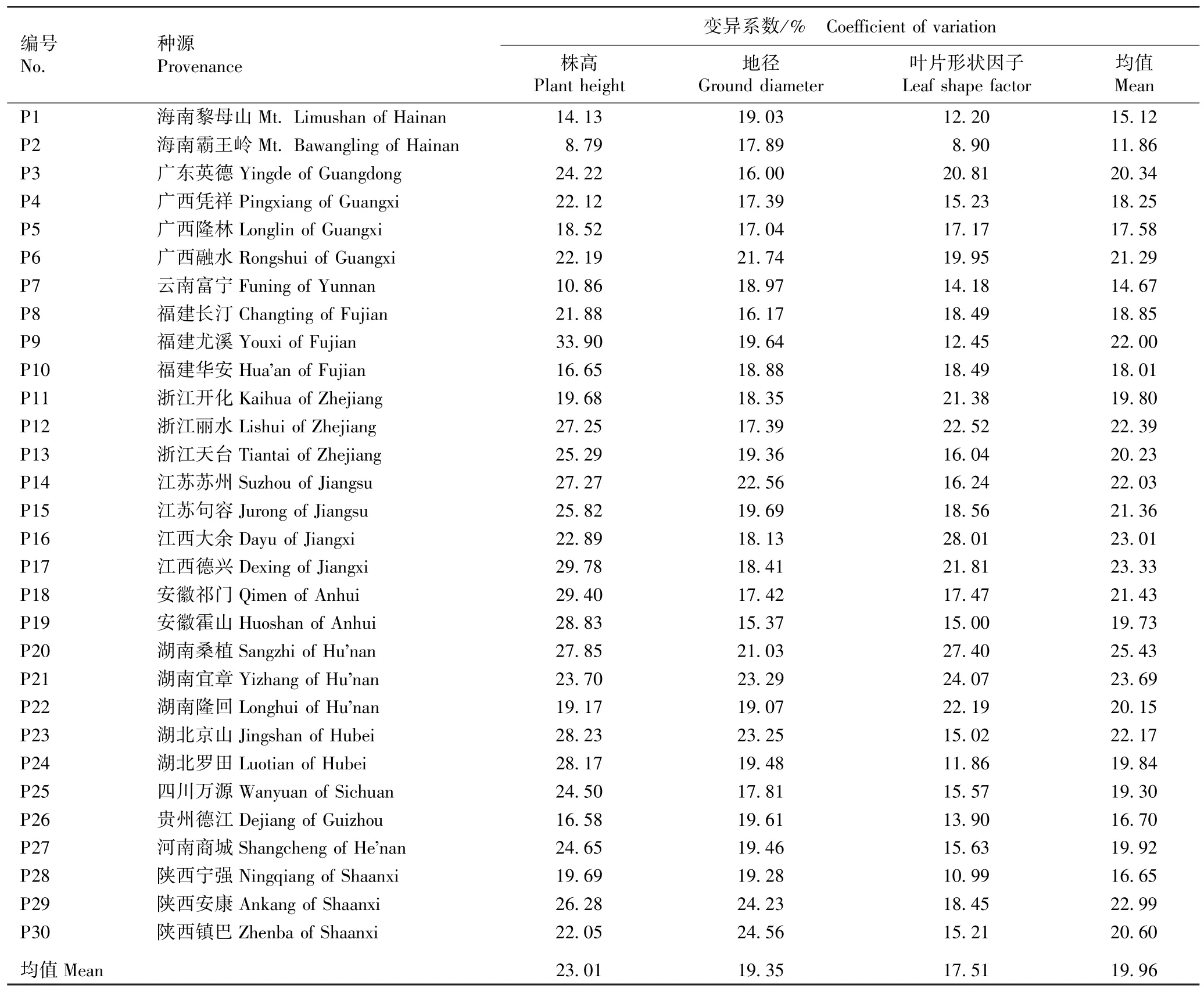
表2 不同种源枫香树幼苗生长性状变异系数的比较Table 2 Comparison on coefficients of variation of growth traits of Liquidambar formosana Hance seedlings from different provenances

表3 枫香树幼苗生长性状与其种源地地理-气候因子间的相关系数Table 3 Correlation coefficient between growth traits of Liquidambar formosana Hance seedlings and geographical-climatic factors of their provenance locations
2.4 逐步回归分析
地理-气候因子对枫香树幼苗株高、地径和叶片形状因子的影响存在共线关系,剔除逐步回归过程中存在共线关系的地理-气候因子,结果(表4)显示:株高与无霜期和海拔存在极显著的线性关系,地径和叶片形状因子与1 月均温也存在极显著的线性关系,这3 个地理-气候因子能解释枫香树幼苗生长性状30.8%~81.9%的变化。

表4 枫香树幼苗生长性状与地理-气候因子的逐步回归分析结果Table 4 Result of stepwise regression analysis on growth traits of Liquidambar formosana Hance seedlings and geographical-climatic factors
3 讨论和结论
3.1 枫香树苗期性状变异分析
株高和地径是研究林木苗期生长的重要指标,在香椿〔Toonasinensis(A. Juss.) Roem.〕[18]、刨花楠(MachiluspauhoiKanehira)[19]和青钱柳〔Cyclocarya paliurus(Batal.) Iljinsk.〕[20]等苗期研究中均作为生长差异的重要参考,其中,香椿苗高和地径的变异系数分别为38.58%和14.19%,刨花楠苗高和地径的变异系数分别为8.60%和5.75%,而本研究中,枫香树幼苗株高和地径的变异系数分别为23.01%和19.35%,说明株高的变异普遍较地径丰富,且枫香树幼苗株高的变异水平居中,地径的变异水平相对较高。 叶片形状因子是综合描述叶面积与叶片周长的指标,也是叶片生长的重要指标。 枫香树幼苗叶片形状因子的变异系数为17.51%,较株高和地径的变异系 数 小, 但 较 红 榉(ZelkovaschneiderianaHand.-Mazz.)[21]叶片形状因子种源间平均变异系数(9.34%)大。 枫香树幼苗的株高、地径和叶片形状因子在种源间存在明显差异,这种差异可以作为区别不同种源的依据。 本研究未对枫香树种源内家系间子代苗期生长性状的差异进行挖掘,关于其生长性状的变异主要来源于种源间还是家系间还需进一步研究。由于本实验是在同一地块的相近区域展开,且后期人工管理措施一致,因此,可初步认定这些生长差异主要受遗传影响。
3.2 枫香树的生长节律与速生种质选择
相关研究结果表明:红椿[7]、台湾桤木〔Alnus formosana(Burkill) Makino〕[22]和任豆(ZeniainsignisChun)[23]的生长节律均符合Logistic 方程拟合模型(R2>0.95),红椿和任豆苗高和地径的生长均呈现“慢—快—慢”的“S”型生长曲线模式,可将苗高和地径的生长分为生长前期、速生期及生长后期3 个阶段;台湾桤木年生长进程划分为渐增期、速生期和缓增期3 个阶段,其中处于速生期植株的株高和地径生长量均超过全年总量的70%。 南方红豆杉〔Taxuswallichianavar.mairei(Lemée et Lévl.) Cheng et L.K. Fu〕[24]的苗高、地径和苗高生长节律具有较丰富的遗传差异,遗传改良潜力较大,早期选择有一定的可靠性。 本研究中,枫香树幼苗的株高和地径存在2 个快速增长阶段,且部分种源株高在4 月至8 月的生长速度较12 月至次年3 月更快。 刘军等[18]对香椿的研究结果表明:生长越快,抗冻性越差。 30 个种源中,云南富宁、海南霸王岭和海南黎母山种源枫香树幼苗的株高生长迅速,可作为速生种质进行进一步研究。 这3 个枫香树种源分布地的平均温度相对较高,其是否适应于偏北地区,还需要进行抗冻性方面的研究。 相比较而言,陕西宁强和陕西镇巴种源枫香树幼苗的株高和地径生长居中,可作为耐寒速生种质进行进一步研究。
3.3 枫香树苗期性状地理变异分析
广布植物种群地理变异的形成与自然选择、基因流和基因飘动等因子有关。 植物的变异形式有连续变异、区域板块变异以及随机变异等模式[25]。 核桃楸(JuglansmandshuricaMaxim.)[26]果核性状变异是不连续变异;而花楸树[2]表型性状为纬度和经度双向变异模式,其中纬度起主要作用;与枫香树分布区相似的樟树〔Cinnamomumcamphora( Linn.)Presl〕[27]的种子长宽比、杉木〔Cunninghamia lanceolata(Lamb.) Hook.〕[28]的株高和胸径以及香椿[18]的苗高、地径和抗冻性是以纬度变异为主;红椿[7]的苗高和地径以经度变异为主。 相关性分析结果显示:枫香树幼苗的株高与种源地的纬度和经度极显著相关,而地径和叶片形状因子仅与经度极显著相关,这种影响表现在不同种源幼苗生长性状差异上,并形成了在地理空间上株高呈“西南—东北”递减、地径呈“南—北”递增和叶片形状因子呈“北—南”递减的变化趋势。 逐步回归分析结果显示:枫香树幼苗的株高主要受无霜期和海拔影响,地径和叶片形状因子主要受1 月均温影响,集中表现为由种源地温度差异引起的种源间生长性状差异,且除株高与海拔存在线性关系,其他2 个生长性状与海拔没有直接的线性关系。 除了纬度对气温有影响外,海拔也会引起枫香树分布区内的温度差异,因此,在枫香树优树选择过程中,既要考虑种源的纬度变异模式,也要考虑海拔引起的温度变化所造成的种源差异。 本文没有进一步根据苗期生长性状进行聚类分析,只进行了单个性状的地理空间变化分析。 由枫香树3 个生长性状的地理变化趋势可以发现,三者间并没有统一的变化规律。 部分相邻枫香树种源幼苗的株高、地径和叶片形状因子间存在显著差异,表明在一定范围内枫香树种源间苗期生长性状的变异是随机的。 在进一步讨论枫香树种源区划时,需要考虑育种目标性状的选择。综上所述,30 个枫香树种源1 年生幼苗生长性状存在丰富变异,这对速生耐寒枫香树种质的选择具有理论指导意义。
