高温和干旱胁迫对西红柿幼苗生长、养分含量及元素利用效率的影响
2019-06-04王德福段洪浪黄国敏周际海黄荣珍樊后保
王德福,段洪浪,黄国敏,周际海,李 威,黄荣珍,樊后保
南昌工程学院江西省退化生态系统修复与流域生态水文重点实验室, 南昌 330099
随着全球气候的变暖,预计到本世纪末,全球平均气温最高可升高6℃[1]。同时未来发生极端气候事件(高温、极端干旱)的频度与强度都会增加[2]。与逐渐变化的气候现象相比,极端气候变化可能会对植物产生更大影响[3-4]。有研究表明高温与干旱经常同时发生[5],其中一种因子的出现往往会加剧另外一种因子的影响程度,进而对植物的生长产生更大的消极影响[6-7]。因此,研究高温与干旱交互作用对农作物的影响将为准确评价农作物对未来极端气候条件的响应提供科学依据。
极端气候现象(高温、干旱)的出现将对植物的生理生态产生一定消极影响,甚至导致植物死亡,但是植物可通过自身的调节在一定程度上来抵抗这些不利影响[8]。高温胁迫主要通过诱导光系统Ⅱ活性中心转化为无活性中心,从而降低植物的光合速率[9],抑制植物生长,抑制生物量的积累。干旱胁迫会破坏植物的叶绿体结构,抑制与光合有关酶的表达,对植物的生长产生不利影响[10-11]。同时有研究表明,温度与水分会改变植物对营养物质的利用,影响植物的生物量分配[12],也会对植物体养分含量产生影响[13]。温度升高会导致植物叶片的氮、磷含量降低[14-15]。干旱会使植物对氮、磷吸收的减少[16-17],可能导致植物氮、磷含量减少。虽然干旱经常伴随着高温出现,但有关高温与干旱交互作用对植物生理生态特征的影响仍然具有很大不确定性[18]。
西红柿(Solanumlycopersicum)被认为是全球第四大经济作物,也是种植面积最大、深受广大消费者喜爱的蔬菜之一,通常种植于户外或者温室[19-21]。同时,西红柿起源于亚热带,经历过多变的温度与突然的高温[22],通常被应用于一些高温研究中[23]。但高温与干旱胁迫交互作用对西红柿幼苗生长与生理生态特征的影响还不清楚。本文选取西红柿幼苗作为试验对象,研究高温与干旱胁迫对西红柿幼苗的生物量积累与分配、养分含量、光合元素利用效率的影响,揭示高温、干旱单独作用以及其交互作用对西红柿生长、养分含量及利用效率的影响机制,以期为全球气候变化的背景下西红柿的种植与培育提供科学依据。我们提出如下假设:(1)高温、干旱以及高温干旱胁迫均会降低西红柿幼苗的生物量,并且高温与干旱胁迫具有交互效应;(2)高温胁迫会降低幼苗根冠比,干旱胁迫会提高幼苗的根冠比、降低叶质量分数;(3)高温、干旱胁迫会降低幼苗叶片全氮、全磷含量以及光合氮素利用效率、光合磷素利用效率,而高温在一定程度上会加剧干旱胁迫的影响。
1 材料与方法
1.1 试验材料与设计
2015年4月将西红柿种子进行播种,待长成幼苗后,将幼苗移植到容量为3.7 L的塑料花盆中继续生长1个月(土壤来源于附近人工林,红壤)。选取长势良好(株高15—20 cm)的幼苗24盆,随机放在两个人工智能气候箱(型号HP1000GS,中国武汉产)中。每个气候箱中放入12盆,气候箱中温度控制在26℃/19℃(白天/夜间)(白天15 h,夜间9 h),相对湿度保持在60%,光照为250 μmol m-2s-1。试验开始前,每天对幼苗进行浇水,每周施肥(可溶性营养肥料,施可得园艺肥料有限公司,中国武汉产)一次。每周对两个气候箱中的幼苗进行位置互换,以减少气候箱的环境条件异质性对试验结果造成的影响。
试验设置两个温度梯度,分别为常温(白天/26℃,夜间/19℃)与高温(白天/42℃,夜间/35℃),以及两个水分梯度,分别为正常浇水、干旱。因此本研究共4个处理,即CK(对照)、H(高温、正常浇水)、D(常温、干旱)、HD(高温、干旱),每种处理设置5个重复。试验的第1天到第10天,只对幼苗进行轻度干旱处理,即保持叶片气孔导度在0—0.05 mol m-2s-1,具体操作方法参照Duan等[24]。第11天到第17天,对H与HD处理幼苗进行为期7d的高温胁迫。高温处理方法如下:采用逐步增加温度方式以便幼苗适应温度变化,温度每次增加4℃,从26℃增至42℃为止,增温在1天内完成。7d高温胁迫结束后(即第17天),气候箱温度调为常温。第20天,对D与HD处理的幼苗进行复水,即浇水使土壤含水量达到田间持水量。本试验所有指标均在复水后测定。试验期间每天16:00—17:00对D与HD处理的盆栽进行称重并测定土壤体积含水量(SWC),以保持一致的干旱程度。干旱处理的幼苗SWC保持在25%左右,正常浇水的幼苗SWC保持在60%左右。
1.2 生物量、生物量分配的计算
试验第1与第22天采用全收割法对西红柿进行收割,将幼苗分为根、茎、叶,并将根系的泥土洗干净。然后将样品放入60℃烘箱中烘干48 h至恒重,分别称取根、茎、叶干重(g),并计算出单株幼苗的总生物量(根、茎、叶生物量相加)。同时根据公式计算出根冠比(地下生物量/地上生物量)、根质量分数(根生物量/总生物量)、茎质量分数(茎生物量/总生物量)、叶质量分数(叶生物量/总生物量)。使用LI-3000C便携式叶面积仪(Li-Cor Inc., Lincoln,NE, USA)测定植物叶面积(cm2),通过计算得出比叶面积(叶面积/叶干重,cm2/g)。
1.3 植株全氮(TN)、全磷(TP)含量的测定
分别将烘干的根、茎、叶进行研磨后过60目筛,称取100—200 mg,经H2SO4-H2O2消煮得到待测液。采用TOC仪测定植物全氮,用钼锑抗比色法测定植物全磷。单位用植物单位质量的养分含量表示(mg/g)。根据公式计算出植物各部分氮磷比(全氮含量/全磷含量)。使用LI-6400便携式光合仪(Li-Cor Inc., Lincoln, NE, USA)测定植物叶片光合速率,具体操作方法参照Duan等[7]。计算得出光合氮、磷元素利用效率(PNUE,PPUE)(光合速率/叶片全氮、磷元素含量,μmol CO2g/m2s mg)。
1.4 数据处理
首先利用SPSS 19.0进行双因素(温度和水分)方差分析,如果交互作用显著,则进一步利用单因素方差分析及Duncan法进行处理间的差异显著性检验。所有结果P<0.05即显著。采用SigmaPlot 12.5作图。
2 结果与分析
2.1 高温与干旱胁迫对西红柿幼苗生长的影响
2.1.1对西红柿幼苗生物量积累与比叶面积的影响
高温与干旱均会降低幼苗的生物量,并且高温与干旱表现出一定的交互效应。与对照相比,H处理使总生物量减少43%,其中对根部影响最大,使根部生物量减少57%。D处理使总生物量减少51%,对叶生物量影响最大,使叶生物量降低62%。HD处理对生物量影响显著大于高温与干旱单独作用,使总生物量减少66%。因此,高温、干旱胁迫能够抑制西红柿幼苗的生长,并且高温加剧了干旱对西红柿幼苗生长的影响,同时干旱也加剧了高温的影响。高温、干旱单独作用以及其交互作用均未对比叶面积产生显著影响(表1,表2)。
表1 温度与水分对西红柿幼苗生长及养分含量影响的双因素方差分析
Table 1 Two-way ANOVA on the effects of temperature and water treatments on growth, nutrient concentration and nutrient use efficiency of tomato seedlings

指标Parameters温度Temperature水分Water温度×水分Temperature×waterFPFPFP总生物量Total biomass4.4880.2817.3390.2257.8980.016根冠比Root shoot ratio0.2080.7285.1780.2646.8060.023比叶面积Specific leaf area4.1360.2910.6600.4300.6380.439根质量分数Root mass fraction0.2250.7185.0780.2666.9680.022茎质量分数Stem mass fraction2.5290.357160.7870.0500.4180.530叶质量分数Leaf mass fraction4.6540.2761405.0910.0170.0580.814叶全氮含量Leaf total nitrogen concentration2.6720.3506.5560.2372.9910.114茎全氮含量Stem total nitrogen concentration2.5320.3570.2900.6850.5230.484根全氮含量Root total nitrogen concentration0.8400.6562.1570.3815.3610.039叶全磷含量Leaf total phosphorus concentration0.6730.5630.2900.6867.5460.021茎全磷含量Stem total phosphorus concentration0.7000.5568.6130.2099.1720.010叶氮磷比Leaf N/P ratio0.0190.91323.7270.1290.5970.457茎氮磷比Stem N/P ratio0.1520.76319.7320.14111.3440.006光合氮素利用效率Photosynthetic nitrogen use efficiency2.3480.3680.0370.88016.6800.002光合磷素利用效率Photosynthetic phosphorus use efficiency5.2920.2610.2550.70210.2900.008

表2 不同处理对西红柿幼苗生物量及比叶面积影响
表中数值表示为平均值±标准误(Values=Mean±SE),同一列中不同字母表示处理之间存在显著差异(P<0.05); CK:对照,Control; H:高温,High temperature;D:干旱,Drought;HD:高温干旱,High temperature plus drought
2.1.2对西红柿幼苗生物量分配的影响
与对照相比,H处理使根质量分数降低了24%,D处理使根质量分数显著增加22%,HD处理使根质量分数显著增加26%(图1,表1)。另外,干旱(D、HD)处理显著降低了西红柿幼苗的叶质量分数。由表1与图1分析可知,干旱处理显著增加茎质量分数。不同处理对根冠比的影响与对根质量分数的影响类似。进一步分析可知,在生物量分配的影响方面,D与HD处理之间不存在显著差异,然而H与HD存在显著差异,说明与高温相比,干旱对西红柿幼苗生物量分配起主要作用。

图1 不同处理对西红柿幼苗生物量分配的影响(数值=平均值±标准误)Fig.1 Effects of different treatments on biomass allocation of tomato seedlings (Values=Mean±SE,the same below)不同字母表示同一指标在处理之间存在显著差异(P<0.05)下同。CK:对照,Control; H:高温,High temperature;D:干旱,Drought;HD:高温干旱,High temperature plus drought
2.2 高温与干旱对西红柿幼苗养分含量及元素利用效率的影响
2.2.1对西红柿幼苗养分含量的影响
由图2分析可知,在叶片方面,与对照相比,H处理导致叶片TN含量显著降低(22%),D与HD处理未对叶片TN含量产生显著影响。H、D处理导致叶片TP含量显著降低,D处理显著提高了叶片N/P。由图2分析可知,与对照相比,H、D、HD处理均未对茎TN含量产生影响,而H处理导致茎TP含量显著提高,D处理导致茎TP含量显著降低。H处理下茎的N/P显著降低,而D处理显著提高了茎N/P。与对照相比,H处理未对根TN含量产生影响,D处理导致根TN含量显著提高,H与HD处理导致茎TP含量显著降低。H与HD处理下根的N/P显著提高(图2)。分析发现,高温未加剧干旱对幼苗叶、茎、根的TN、TP含量以及N/P的影响。
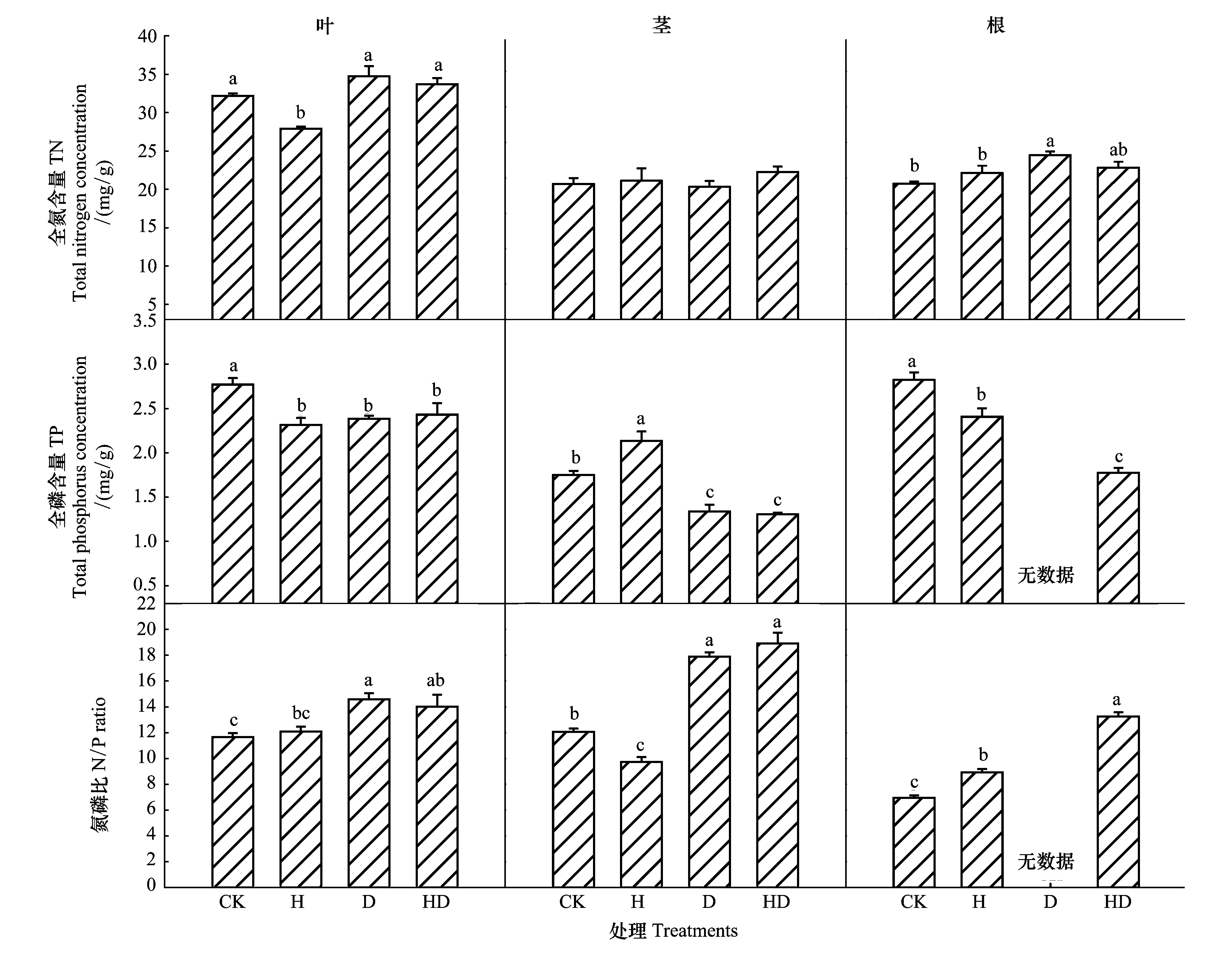
图2 不同处理对西红柿幼苗元素含量的影响Fig.2 Effects of different treatments on nutrient concentration of tomato seedlings
2.2.2对西红柿幼苗光合元素利用效率的影响
与对照相比,H处理使PNUE降低49%,D处理使PNUE降低30%,HD处理使PNUE降低25%。D与HD对PNUE的影响不存在显著差异。与对照相比,H使PPUE降低48%, D对PPUE未产生显著影响,而HD使PPUE降低26%并显著低于D处理(图3,表1)。

图3 不同处理对西红柿幼苗光合元素利用效率的影响Fig.3 Effects of different treatments on photosynthetic nutrient use efficiency of tomato seedlings
3 讨论
3.1 高温与干旱对幼苗生长特性的影响
3.1.1高温与干旱对生物量的影响
生物量是植物生长状态的综合反映,本试验的研究结果与假设(1)基本相同。与对照相比,H处理胁迫能显著降低西红柿幼苗的根、茎、叶生物量以及总生物量。有研究结果显示,在高温胁迫条件下,五个品种的西红柿的生物量均显著降低[25]。同时,有研究也得出结论,极端高温(42℃)处理均能显著降低西红柿地上部分与地下部分的生物量[26]。干旱胁迫也会使植物的生长受到抑制,降低植物总生物量的积累[27]。本研究结果与之前的研究结果一致,与对照相比,D处理能显著降低幼苗总生物量以及跟、茎、叶生物量。有研究也得出类似结论,与对照(正常浇水)比较,干旱处理均能使水稻(Oryzasativa)的茎、叶与总生物量显著降低[28]。此外,无论是中度干旱(田间持水量的50%)还是重度干旱(田间持水量的35%)均能显著降低玉米(Zeamays)总生物量[29],而且植物生物量的降低程度随着干旱程度的增加而加剧[30-31]。
高温与干旱胁迫的交互作用也会影响植物生物量积累。与对照相比,羊草(Leymuschinensis)生物量在高温与干旱交互作用下显著减少,并且减少量大于高温与干旱单独作用[32]。有研究也得出类似的结论,与对照相比,高温干旱双重作用使玉米的总生物量下降的最多(约75%,对于高温与干旱单独作用)[33]。本试验的结果也表示,与对照相比,HD处理显著降低了西红柿幼苗的总生物量,并且降低的幅度明显高于H与D影响的结果,说明高温与干旱对植物的生长的影响具有一定的交互作用。
3.1.2高温与干旱对生物量分配格局的影响
本研究结果与假设(2)相同。与对照相比,H处理能显著降低西红柿幼苗的地上、地下部分生物量,但对地下部分的影响更大,因而导致根冠比、根质量分数显著降低。有研究表明也表明,高温胁迫下,九种西红柿幼苗的根冠比均显著降低,主要原因是高温对地下部分的生长抑制作用比地上部分大[25]。另外,高温能降低西红柿的地上、地下部分的生物量,但是与地上部分相比,地下部分对温度升高更敏感,进而导致根冠比显著降低[26]。而植物地下部分比地上部分更敏感于温度升高的主要原因是地下部分最佳生长温度低于地上部分[34]。
生物量分配能够反应植物生境的资源情况。根据最优分配理论与功能平衡原则,植物生长受到资源限制时,会增加和优先配置一些能获得该资源的器官或组织以抵御胁迫环境[35]。有研究得出结论,与对照组(正常浇水)相比,干旱处理条件下,水稻会分配更多生物量到地下部分,导致根质量分数与根冠比增加,叶质量分数降低[28]。本研究也表明,与对照对相比,D处理增加了幼苗的根质量分数,显著降低了幼苗叶质量分数,增加了根冠比。这样有利于植物在干旱条件下吸收到更多的水分,减少地上部分的水分散失,从而减少干旱胁迫所造成的伤害[36]。另外有研究发现,植物在干旱条件下会相对增加地下部分的生物量,不仅可以促进根对水分的吸收,也可能会提高根对氮素的吸收,这将使植物有更多的氮投入到光合系统[37-38]。然而干旱胁迫改变生物量分配格局的机理各不相同。有学者研究结果表明,中度干旱处理对玉米的地上部分生物量有显著抑制作用,但对地下部分生物量没有显著影响,从而使根冠比增加,而重度干旱均有显著降低对地上、地下生物量,但地上部分降低更多,进而根冠比增加[29]。也有研究表明干旱胁迫会促进生物量更多的分配到地下部分,减少对地上部分的分配,进而改变生物量分配格局,使根冠比增加[39]。但也有一些研究表明,干旱处理不会对根冠比产生显著影响甚至会使根冠比降低[40-42]。此外,有研究认为,干旱胁迫会显著增加冰草(Agropyronfragile)、Elymuselymoides的根质量分数,明显减少Festucaidahoensis的根质量分数[43]。综上所述,干旱胁迫对植物生物量分配格局的影响存在一些不确定性,可能与植物品种、干旱时间以及强度有关。
另外,在本研究中,高温与干旱的交互作用对幼苗生物量分配的影响有异于单独作用产生的影响。与对照、H处理相比,HD处理显著增加了幼苗的根冠比、根质量分数、茎质量分数,显著降低了叶质量分数。但与D处理相比,HD处理并没有产生显著变化。说明与高温胁迫相比,干旱在西红柿幼苗生物量分配方面起主导作用。
3.1.3高温与干旱对幼苗比叶面积的影响
比叶面积是植物叶片的重要性状,能够反映植物对不同生存环境的适应特征[44]。有关高温对植物的比叶面积的影响存在异议。有研究表明,白天高温会导致豆类作物的比叶面积降低,夜晚高温会使比叶面积增加(叶面积没有什么变化),使比叶面积增加的可能原因是夜晚的呼吸作用降低了叶片生物量,进而使比叶面积增加[45]。有研究表明,干旱处理会降低植物的比叶面积[32,46]。本研究结果表明,与对照相比,H、D、HD处理均未对幼苗比叶面积产生显著影响,可能原因是经过长期进化,西红柿幼苗已经适应了高温干旱环境。
3.2 高温与干旱对幼苗养分含量的影响
植物的生长需要很多种元素,其中包括大量元素(C、N、P等)。温度的变化会影响植物体内酶的活性、反应介质等来调节植物细胞功能的发挥,影响植物N、P等元素代谢,进而影响N、P含量以及N/P[47]。同时叶片N/P能够反映植物所生环境的氮磷限制状态[48]。当前普遍采用的是Koerselman和Meuleman[49]的结论,即N/P<14表明植物受到N限制,N/P>16表明植物受到P限制。有研究发现,温度升高能降低4种森林类型植物叶片的N含量、增加叶P含量、N/P降低[14]。另外有研究表明,随着温度升高导致陆生植物叶片的N、P含量降低,但对N/P没有产生显著影响,原因是植物在纬度差异较小情况下,叶片N/P能够保持相对稳定[15]。本研究结果与假设(3)基本类似。与对照相比,H处理导致叶片的TN、TP含量显著降低,而对N/P(12.09<14)没有产生显著影响。根据Koerselman和Meuleman理论推断,幼苗在对照、H条件下均受到N限制。可能原因是红壤氮素含量较低[50],高温胁迫未改变红壤的氮素缺乏状况。此外,与对照相比,H处理未导致茎TN含量产生显著影响,而使茎TP含量显著提高,因而引起茎N/P显著降低。也有研究表明,极端高温胁迫下,植物的根部代谢速率、代谢水平以及养分吸收蛋白的活性均会降低,进而引起根部对养分吸收的减少[26],可能导致根部养分含量的降低。本研究中也得出类似结论,与对照相比,H处理导致根部TP含量显著降低。
水分会影响土壤养分有效性,从而影响植物的N、P含量以及计量比[51]。一些研究表明,干旱胁迫会降低植物对氮、磷元素的吸收、运输、再分配[16]。土壤含水率的下降也会导致植物对氮、磷吸收的减少[17,52]。与植物生长相比,干旱对植物氮、磷吸收产生更大影响,导致氮、磷含量的降低[53]。但干旱条件下复水(土壤水势的快速升高)会导致土壤微生物细胞的裂解[54],释放细胞中的溶质[55],增加了有机物的分解[56]。复水后有机物分解的提高将增加土壤总的以及净氮矿化速率[57],使土壤中有效氮的含量增加(相对于复水之前),植物将吸收到更多的氮,因此,不会对植物氮含量产生影响[53]。本研究得出类似结论,与对照相比,干旱处理(D、HD)未对叶片、茎、根(D处理除外)的TN含量产生显著影响。说明旱后复水增加了本试验中土壤有效氮含量,西红柿幼苗从土壤吸收到更多的氮素。有研究表明,干旱后复水能增加土壤中有效磷含量[58],增加的有效磷量可能弥补了之前干旱条件下有效磷的减少量,不对植物磷含量产生影响[53]。然而在本研究中,与对照相比,旱后复水条件下叶片、茎、根的TP含量均显著降低。原因可能是复水使土壤中有效磷的增加量不够,土壤中有效磷含量仍显著低于对照水平,导致幼苗叶、茎、根磷含量显著比对照水平低。与对照相比,干旱处理(D、HD)显著提高了叶片、茎、根(D处理除外)的N/P(N/P>14),主要原因是干旱处理对叶片、茎、根TN含量没有显著影响,却显著降低了叶片、茎、根TP含量。根据Koerselman和Meuleman理论,说明D、HD缓解了幼苗氮素限制状况。可能是因为复水后土壤中有效氮含量迅速提高[53],改善了幼苗的氮磷含量不均衡状况。
本文研究也表明,与H处理相比,HD处理导致叶片TN含量显著提高,而未对茎、根TN含量产生影响。说明在高温环境下,干旱胁迫有助于西红柿幼苗叶片TN含量的提高。有研究表明,高温干旱胁迫能提高刺槐幼苗体内N的积累[59]。在本研究中,与H处理相比,HD处理导致茎、根TP元素含量显著降低,而未对茎、根TN元素含量产生显著影响,进而导致茎、根的N/P显著提高。同时也表明干旱削弱了了高温条件下西红柿幼苗根部对P元素的吸收。与D处理相比,HD处理均未对叶、茎、根TN、TP含量产生显著影。说明与高温相比,干旱在西红柿幼苗的养分含量方面发挥着主要作用。
3.3 高温与干旱对幼苗光合元素利用效率的影响
光合氮素利用效率指的是单位质量的叶片氮所获取和同化的碳量[60]。本研究表明,与对照相比,H处理显著降低了光合氮素利用效率(PNUE),原因是H处理导致叶片N含量显著降低,但H处理使幼苗叶片的光合速率降低的更多[7]。本研究结果与假设(3)基本一致。与对照相比,D、HD处理均显著降低了PNUE,原因是D、HD处理未导致叶片N含量产生显著变化,却降低了幼苗叶片的光合速率,导致PNUE降低。有研究表明,植物PNUE与植物生长速度以及生物量的积累在一定程度上成正比关系[61]。本文的研究结果显示,与对照相比,高温、干旱与高温干旱交互作用均显著降低了幼苗的生长速率与生物量,H、D、HD均显著降低了幼苗的氮素利用效率,在一定程度上也证实了光合氮素利用效率与生物量积累成正比关系。本研究中,高温、干旱以及高温干旱交互作用对光合磷素利用效率(PPUE)的影响与PNUE类似。同时,PNUE也被认为是一种用来描述叶片生理与资源利策略关系的一种重要的叶片特征[62]。有研究表明,干旱胁迫显著降低了五角枫苗(Acermono)的光合氮素利用效率,主要原因的植物在干旱条件下水分利用效率的提高[63-64]。因此,在全球气候变化背景下,研究高温与干旱胁迫对西红柿幼苗的光合元素利用效率的影响将有助于西红柿种植、培育过程中的水肥管理。
4 结论

图4 干旱条件下,高温胁迫对西红柿幼苗生长与养分含量的影响Fig.4 Effects of high temperature on growth, nutrient concentration and nutrient use efficiency of tomato seedlings under drought stress图中的↓表示显著降低, —表示没有显著影响
本研究主要集中于高温、干旱单独作用以及交互作用对西红柿幼苗的生长、养分含量及利用效率的影响。研究结果表明,高温、干旱单独作用以及交互作用均显著降低了西红柿幼苗总生物量的积累,并且高温与干旱对生物量积累的影响具有一定的交互效应。与对照相比,高温单独作用显著降低了幼苗根冠比与根质量分数,干旱单独作用显著提高了幼苗根冠与根质量分数、显著降低了叶质量分数。高温未加剧干旱胁迫对幼苗生物量分配的影响。高温、干旱单独作用以及交互作用均未对幼苗比叶面积产生显著影响。与对照相比,高温单独作用显著降低了叶片TN、TP含量、提高了茎TP含量、降低了根TP含量。干旱单独作用显著提高了根TN含量、显著降低了叶、茎、根的TP含量。高温未加剧干旱胁迫对幼苗养分含量的影响。在光合元素利用效率方面,与对照相比,高温、干旱单独作用均降低了幼苗PNUE、PPUE,并且高温加剧了干旱胁迫对幼苗光合磷素效率的影响。
综上所述,表明:(1)高温在一定程度上加剧了干旱胁迫对西红柿幼苗生长与光合磷素利用效率的影响;(2)高温未加剧干旱胁迫对幼苗生物量分配与养分含量的影响(图4)。
