利用荧光SSR分析中国糜子的遗传多样性和群体遗传结构
2019-05-28寇淑君霍阿红付国庆纪军建王瑶左振兴刘敏轩陆平
寇淑君,霍阿红,付国庆,纪军建,王瑶,左振兴,刘敏轩,陆平
(1张家口市农业科学院,河北张家口075000;2中国农业科学院作物科学研究所,北京 100081)
0 引言
【研究意义】糜子(Panicum miliaceumL.)属于禾本科黍属(Panicum),为一年生草本植物,有2n=4x=36条染色体。糜子起源于中国,是最古老的栽培作物,距今已有10 000余年的栽培历史[1],具有抗旱、耐盐碱、耐瘠,生育期短等特点[2],不仅为干旱半干旱地区的人们提供粮食保障,也是该地区重要的救灾备荒作物[3]。中国糜子种质资源非常丰富,目前,保存在国家种质库中的糜子资源超过8 800多份,如何准确快速评估所保存资源的遗传多样性和群体遗传结构,对于开展糜子抗逆相关基因的挖掘,促进糜子种质创新和品种改良等具有重要意义。【前人研究进展】分子标记是揭示不同品种间遗传多样性和亲缘关系的有效手段[4],RAPD、AFLP等分子标记曾被用于糜子的遗传多样性分析。M'RIBU等[5]应用RAPD技术分析了 4类糜子的遗传多样性;KARAM 等[6]用AFLP标记检测了12份来自美国和加拿大的糜子遗传多样性;LÁGLER等[7]使用来自草莓属植物的 9个ISSR标记在21份匈牙利糜子中共检测到15个等位基因,其中7个引物能扩增出DNA片段。SSR分子标记具有共显性、在基因组中含量丰富、多态性信息高、重复性好等优点[8],被广泛用于各种作物的遗传多样性研究中[9-12]。近年来,利用SSR分子标记对糜子的遗传多样性研究也取得了一些进展。HU等[2]用46对来源于水稻、小麦、燕麦和大麦的SSR标记分析了118份糜子资源的遗传多样性,PIC值为 0.284—0.980。CHO等[13]基于糜子基因组DNA富集SSR文库开发了首批糜子SSR标记,利用25个标记在50份材料中检测到110个等位变异。HUNT等[14]用16个标记检测了 98份来源于欧亚大陆的糜子地方品种的遗传多样性。连帅等[15]、LIU等[16]、薛延桃等[17]、王银月等[18]用高通量测序手段开发的SSR标记全面分析了中国糜子地方品种、育成品种、野生材料以及新引进国外资源的遗传多样性和群体遗传结构。与传统的聚丙烯酰胺凝胶电泳相比,毛细管电泳检测技术具有灵敏度高、分辨率高、重复性好、所需样品量少、分析通量大、可以有效减少错读等优点[19],在玉米[20]、小麦[21]、烟草[22]、油菜[23]等植物中广泛应用,但在糜子上相关报道较少[14,24]。【本研究切入点】已开展的遗传多样性研究结果显示,糜子虽然是异源四倍体作物,但是其遗传多样性水平总体不高,一方面可能是目前可用于糜子遗传多样性研究的SSR分子标记较少且多态性不高;另一方面多数研究采用的传统聚丙烯酰胺凝胶电泳检测技术分辨率较低,不能准确反映糜子不同材料间的遗传多样性。【拟解决的关键问题】本研究选用严格筛选后的22对SSR引物进行荧光标记,利用全自动基因分析仪检测技术分析来自中国不同生态区的72份糜子育成品种及59份当地主要农家种的遗传多样性和群体遗传结构,为糜子品种改良、种质创新和资源有效利用提供依据。
1 材料与方法
1.1 试验材料
试验材料包括由国家种质资源库提供的 72份育成品种(宁夏回族自治区11份、辽宁省2份、吉林省8份、黑龙江省10份、甘肃省8份、山西省16份、内蒙古自治区14份和陕西省3份)和59份农家种(宁夏回族自治区4份、辽宁省2份、吉林省2份、黑龙江省6份、甘肃省4份、山西省10份、内蒙古自治区5份、陕西省5份、青海省8份、河北省9份、新疆维吾尔自治区3份和山东省1份)(电子附表1)。根据糜子栽培生态区划[25],131份材料所属生态区包括东北春糜子区(30)、华北夏糜子区(3)、北方春糜子区(59)、黄土高原春夏糜子区(36)和西北春夏糜子区(3)。
1.2 DNA提取与检测
每份材料随机选取10个单株,采集三叶一心期幼嫩叶片混合后液氮中研磨,用新型植物基因组 DNA快速提取试剂盒(北京鼎国昌盛生物有限公司)提取基因组DNA,用1%琼脂糖凝胶电泳检测DNA完整性,再用NanoDropND-1000检测浓度并将扩增浓度调成 50 ng·μL-1。
1.3 SSR引物
根据中国农业科学院作物科学研究所小宗作物课题组通过高通量测序技术开发的糜子基因组SSR引物和文献中发表的引物信息,共选取202对SSR引物进行合成。其中22对来源于连帅等[16],4对来源于王银月等[18],178对来源于未发表的高通量测序开发的糜子SSR引物。对筛选出的SSR标记的正向引物5′末端用6-FAM、HEX、ROX和TAMRA 4种荧光染料中的一种进行标记。引物的合成和标记均由英潍捷基(上海)贸易有限公司完成。
1.4 PCR扩增与标记检测
1.4.1 PCR扩增 PCR扩增体系包含11 μL的反应体积,其中PCR Mix(北京擎科生物技术有限公司)5.5 μL、正向引物和反向引物各0.6 μL、基因组DNA 1 μL和ddH2O 3.3 μL。首先通过温度梯度PCR扩增,确定引物的最适退火温度。PCR反应程序(以退火温度55℃为例)为98℃ 3 min;98℃ 10 s,55℃ 10 s,72℃ 10 s,30个循环;72℃ 3 min,4℃保存。
1.4.2 变性聚丙烯酰胺凝胶电泳检测 6%的聚丙烯酰胺凝胶,恒功率65 W预电泳15—20 min,每个加样孔点入3 μL样品,65 W恒功率电泳1—1.5 h,银染检测扩增产物。
1.4.3 毛细管电泳荧光检测 将6-FAM和HEX荧光标记以及TAMRA和ROX荧光标记的PCR产物分别用超纯水稀释10—30倍和5—10倍,随后取等体积的上述4种稀释液混合,从中吸取1 μL加到DNA分析仪深孔板中,同时加入0.1 μL LIZ500分子量内标和8.9 μL去离子甲酰胺。将样品在PCR仪上95℃变性5 min,立即取出置于碎冰上冷却10 min后,放置到ABI 3730 XL型DNA分析仪上进行自动荧光检测。
1.5 数据统计与分析
用Genemapper 4.0软件自动生成每个位点的图谱文件,分析扩增片段峰值,读取扩增片段大小;用PowerMarker3.25[26]计算每对引物的等位基因数(allele number)、主要等位基因频率(major allele frequency)、基因多样性指数(gene diversity index,H)、多态性信息含量指数(polymorphism information content,PIC);用 Popgen1.32[27]计算 Shannon 信息指数(Shannon's Information index,I);在MEGA 6.06[28]中构建遗传聚类图;用STRUCTURE2.2.3[29]软件进行群体结构分析。
2 结果
2.1 引物筛选与验证
利用地理来源远、形态差异大的6份糜子材料对收集的202对SSR引物进行PCR扩增,并对扩增产物进行变性聚丙烯酰胺凝胶电泳检测,淘汰不具多态性或多态性差的引物,筛选出26对条带清晰、多态性较高、扩增稳定的SSR引物。对26对高多态性SSR引物进行毛细管荧光电泳检测,结果表明,其中有22对引物的峰图简单、多态性高、等位变异大小变化符合规律,可用于全部参试材料的遗传多样性分析(表1)。入选的22对引物既适用于传统聚丙烯酰胺凝胶电泳检测技术,也适用于荧光 SSR标记-全自动分析检测技术。
为了检验荧光标记毛细管电泳检测方法的可靠性,用变性聚丙烯酰胺凝胶电泳检测方法对毛细管电泳检测指纹数据进行了验证。以引物BM2654为例(图1),结果显示在BM2654标记位点上,6%变性聚丙烯酰胺凝胶电泳检测到6个等位基因,与图2荧光毛细管电泳检测到的大小分别是197、200、203、206、209和212 bp的多态性片段长度一一对应,说明荧光标记毛细管电泳检测方法结果可靠,能有效用于后续参试材料的遗传多样性研究。

图1 45份糜子材料在BM2654标记位点上的带型表现Fig. 1 Alleles of 45 broomcorn millet accessions at marker locus BM2654

图2 7份糜子材料在荧光标记BM2654位点上的等位变异峰图Fig. 2 Peak patterns detected in seven broomcorn millet accessions at fluorescent marker locus BM2654
2.2 中国糜子种质资源SSR位点多样性分析
利用22对引物在131份材料中共检测出128个主要等位变异,变幅为2—8个,平均每对SSR引物扩增出 5.82个主要等位变异,其中引物 LMX836、LMX2019、LMX2068、LMX2382扩增出的等位变异最多,为8个。主要等位基因频率为0.3015—0.7672,平均0.5241。多态性信息含量(PIC)的变幅介于0.2934—0.8150,平均0.5890,PIC最大的标记为LMX2382,最小的标记为 BM6573。15个位点的 PIC>0.5,为高度多态性位点,7个位点的PIC为0.25—0.5,为中度多态性位点。基因多样性指数的变化范围是 0.3572—0.8132,平均0.6276,其中,BM6573最低,LMX2019最高。Shannon多样性指数为 0.5427—1.7681,平均1.2062,最高为 LMX2019,最低为 BM6573。基因多样性指数,多态性信息含量,Shannon信息指数呈正相关,能够反映每对标记的鉴别能力。22个SSR位点的多态性丰富,可以检测到不同糜子材料间丰富的遗传多样性。

表1 22对SSR标记的遗传参数Table 1 Genetic parameters of the 22 SSR markers used in this study
同时利用这 22对引物分析了不同生态区不同类型糜子资源的遗传多样性(表 2),不同生态区育成品种的基因多样性指数、多态性信息含量、Shannon信息指数的变化范围分别是 0.5474—0.5808、0.4953—0.5337、0.9848—1.1074,平均值为0.5608、0.5111和1.0433,其中黄土高原春夏糜子区育成品种的遗传参数最高,北方春糜子区次之。不同生态区农家种的基因多样性指数、多态性信息含量、Shannon信息指数的变化范围分别是0.3838—0.6221、0.3197—0.5725和0.4402—1.1114,平均值为 0.5041、0.4525和0.7806,其中北方春糜子区农家种的遗传参数最高,黄土高原春夏糜子区次之。总体来讲,北方春糜子区各项遗传参数均最高,黄土高原春夏糜子区次之,华北夏糜子区各项遗传参数均最低,说明北方春糜子区和黄土高原春夏糜子区的遗传多样性比较丰富,北方春糜子区农家种的遗传多样性丰富,可以从中挖掘优良基因用于品种改良。
2.3 中国糜子种质资源的遗传相似性分析
利用 Popgene1.32计算不同生态区糜子种质间的遗传相似性(表3),结果显示,遗传距离的变化范围为0.0764—0.7251,平均0.3121,遗传一致度的变化范围为0.4843—0.9265,平均0.7465。其中,北方春糜子区和黄土高原春夏糜子区的遗传距离最小(0.0764),遗传一致度最大(0.9265),这两个生态区的地理位置相邻,生态环境差异较小,亲缘关系较近。华北夏糜子区和西北春夏糜子区的遗传距离最大(0.7251),遗传一致度最小(0.4843),这两个生态区的地理分布较远,生态环境差异大,亲缘关系较远。说明遗传相似性与地理来源密切相关,地理来源越近,遗传距离越小,遗传一致度越高,亲缘关系越近。
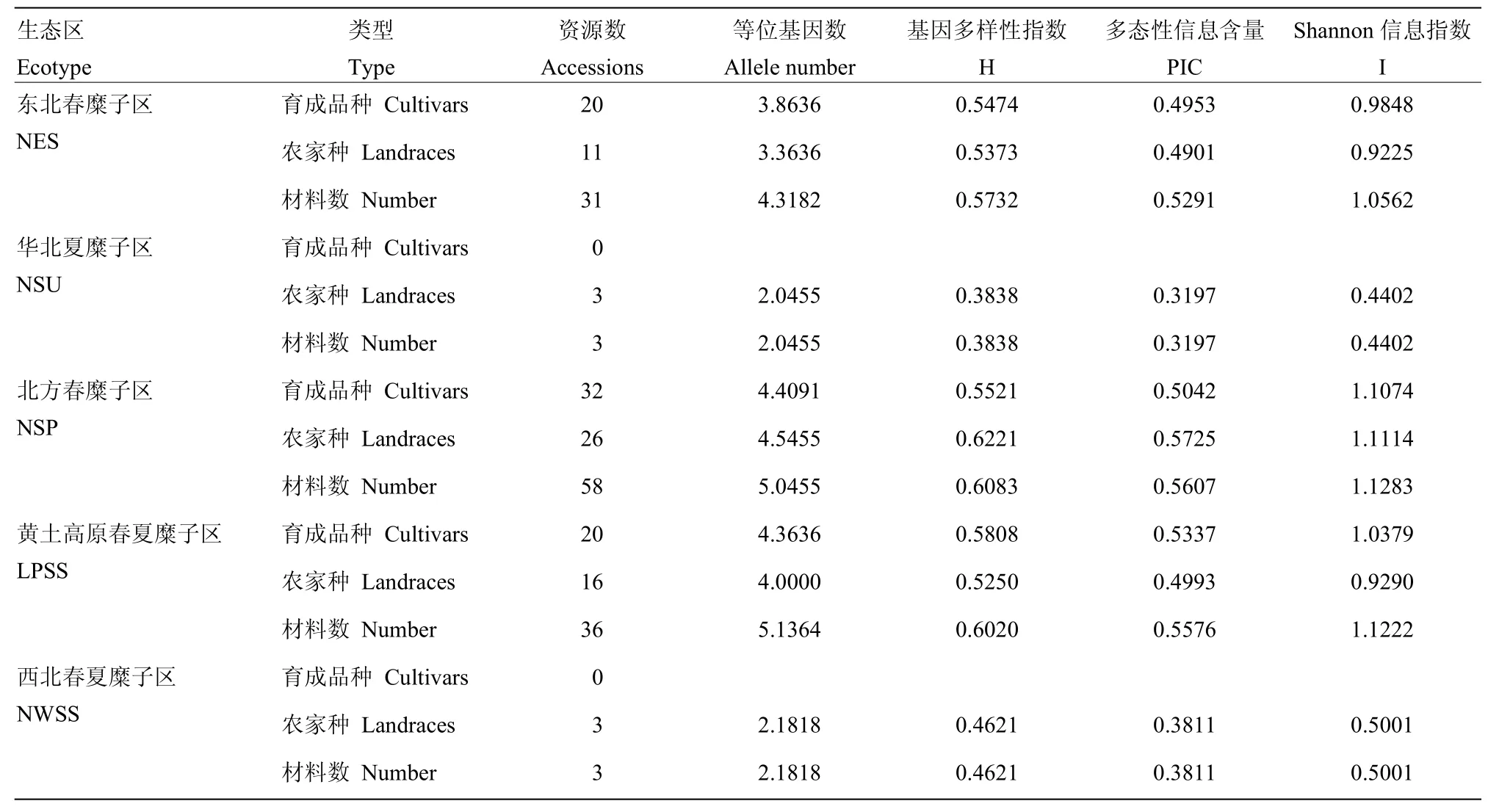
表2 不同生态区糜子的遗传多样性分析Table 2 Evaluation of genetic diversity of different ecotypes of broomcorn millet

表3 不同生态区糜子资源的遗传距离和遗传一致度Table 3 Genetic distance and genetic identity of broomcorn millet accessions with different ecotypes
2.4 基于遗传距离的中国糜子种质资源聚类分析
基于 UPGMA对不同生态区糜子材料进行聚类(图3),在遗传距离0.1284处将5个生态区划分为4个组群(组群Ⅰ、Ⅱ、Ⅲ和Ⅳ)。组群Ⅰ将华北夏糜子区归为一类,材料来自于山东省、河北省。组群Ⅱ将西北春夏糜子区归为一类,材料来自于新疆维吾尔自治区。组群Ⅲ将东北春糜子区归为一类,材料来自于黑龙江省、吉林省、辽宁省。组群Ⅳ将北方春糜子区和黄土高原春夏糜子区归为一类,材料来自于宁夏回族自治区、山西省、陕西省、内蒙古自治区、甘肃省、青海省。聚类结果与遗传相似性分析结果一致。

图3 基于遗传距离的不同生态区糜子资源聚类图Fig. 3 Cluster diagram of broomcorn millet accessions with different ecotypes based on genetic distance
基于 UPGMA对 131份糜子材料进行聚类分析(图4),131份糜子材料聚为7个组群。组群A有30份材料,包括黑龙江省12份、吉林省4份、辽宁省1份、甘肃省2份、山西省3份、陕西省1份、宁夏回族自治区1份、青海省6份,大部分基因型属于东北春糜子区。组群B有7份材料,其中河北省2份、内蒙古自治区2份、宁夏回族自治区、山东省、陕西省各1份,主要属于华北夏糜子区,组群C有3份材料,包括来自山西省的品黍1号,辽宁省的本溪褐糜子和山西省的灰脸蛋糜,表明这3份材料遗传背景相似且与其他材料的遗传差异较大。组群D包括16份材料,其中内蒙古自治区5份、河北省7份、山西省3份、青海省1份,均来自北方春糜子区。组群E包含3份材料,其中内蒙古自治区2份,陕西省1份。组群F有21份材料,包括山西省5份、陕西省4份、甘肃省2份、新疆维吾尔自治区2份,宁夏回族自治区2份、辽宁省1份、吉林省5份,主要属于黄土高原春夏糜子区。组群G包含51份材料,包括山西省11份,内蒙古自治区11份,甘肃省8份,宁夏回族自治区11份、黑龙江省4份,新疆维吾尔自治区、青海省、陕西省、吉林省、辽宁省、河北省各1份,主要属于北方春糜子区和黄土高原春夏糜子区。
东北春糜子区的育成品种中,来自黑龙江省的材料主要聚在组群 A中,来自吉林省的材料主要聚在组群F中,东北春糜子区的农家种与育成品种聚类结果一致。北方春糜子区的育成品种主要聚在组群 G中,农家种主要聚在A、D中,在组群G中也零星出现,其中,来自青海省的农家种集中聚在组群 A中。黄土高原春夏糜子区的育成品种聚在组群A和G中,农家种集中聚在组群F中。来自华北夏糜子区的3份材料全部聚在组群B中。来自西北春夏糜子区的3份材料,2份聚在组群F中,1份聚在组群G中。来自北方春糜子区和黄土高原春夏糜子区的育成品种多数聚在组群 G中,说明这两个生态区的部分育成品种遗传背景相似。农家种在各组群中的分布与地理来源密切相关,部分育成品种的遗传背景可能包含其他生态区的基因型,在各组群中的分布没有明显的区域性。

图4 基于荧光SSR分子标记分析的131份糜子材料聚类图Fig. 4 Cluster diagram of 131 broomcorn millet accessions based on data from 22 SSR markers
2.5 中国糜子种质资源的群体遗传结构分析
用STRUCTURE软件分析来自中国不同地区的131份糜子材料,将 K设为 2—10,绘制 K与 ΔK的关系图(图5),K=4时,ΔK最大,因此在K=4的模式下分析糜子品种的遗传结构[30],并计算出每个类群的最大 Q值分布。不同的色块代表不同的类群(图6,表4),类群Ⅰ为红色部分(35份),代表北方春糜子区,主要包含甘肃省 8份、青海省 7份、内蒙古自治区7份、宁夏回族自治区6份、山西省3份、黑龙江省3份、陕西省1份。类群Ⅱ为绿色部分(21份),包括黑龙江省10份、吉林省3份、山西省3份、甘肃省2份、宁夏回族自治区、陕西省和辽宁省各1份,主要来自东北春糜子区。类群Ⅲ为蓝色部分(43份),包括山西省15份、内蒙古自治区9份、河北省6份、宁夏回族自治区4份、陕西省、黑龙江省、辽宁省各2份、吉林省、新疆维吾尔自治区、青海省各1份,主要来自北方春糜子区。群组Ⅳ为黄色部分(32份),包括山西省 6份、陕西省 5份、吉林省 6份、宁夏回族自治区 4份、河北省 3份、甘肃省、新疆维吾尔自治区各2份、辽宁省、内蒙古自治区、黑龙江省、山东省各1份,主要来自黄土高原春夏糜子区。

图5 131份糜子材料的K值与Δk值的关系Fig. 5 Graphical relationship between K and Δk for 131 broomcorn millet accessions

图6 131份糜子材料的群体遗传结构分析Fig. 6 Population genetic structure of 131 broomcorn millet accessions

表4 131份糜子材料在各组群中的分布(K=4)Table 4 Distribution of 131 broomcorn millet accessions based on STRUCTURE analysis(K=4)
黄土高原春夏糜子区的育成品种在组群Ⅰ、Ⅱ、Ⅲ中都有出现,农家种集中分布在组群Ⅳ中。北方春糜子区的育成品种和农家种主要分布在组群Ⅰ和组群Ⅲ中,其中来自青海省的农家种全部分布在组群Ⅰ中,来自河北省北部的农家种分布在组群Ⅲ中,东北春糜子区的育成品种和农家种主要分布在组群Ⅱ和Ⅳ中,其中来自黑龙江省的材料主要分布在组群Ⅱ中,来自吉林省的材料主要分布在组群Ⅳ中,华北夏糜子区的材料均分布在组群Ⅳ中。西北春夏糜子区的材料分布在组群Ⅲ和Ⅳ中。遗传结构分析结果与聚类结果基本一致。
STRUCTURE群体结构分析中,当某一材料在某类群中的Q≥0.6时,则认为该材料血缘关系相对比较单一,否则认为该材料血缘关系来源复杂[9]。131份糜子材料在各个群体中的Q值分布见表5,131份糜子材料中有 107个品种 Q>0.6,占所有供试材料的81.7%,Q>0.8和Q>0.9的品种分别占58%和48.1%,说明各群体中大部分品种亲缘关系比较单一,较少品种含有其他类群的基因成分。品黍1号在4个类群中的Q值分别为0.190、0.011、0.453和0.346;本溪褐糜子在4个类群中的Q值分别为0.074、0.134、0.443和 0.349;灰脸蛋糜在 4个类群中的 Q值分别为0.047、0.004、0.478和0.471,它们的基因都主要来源于类群Ⅲ和类群Ⅳ中,聚类分析也被单独聚为一个小支,说明这三个品种的遗传背景比较相似,与聚类结果一致。
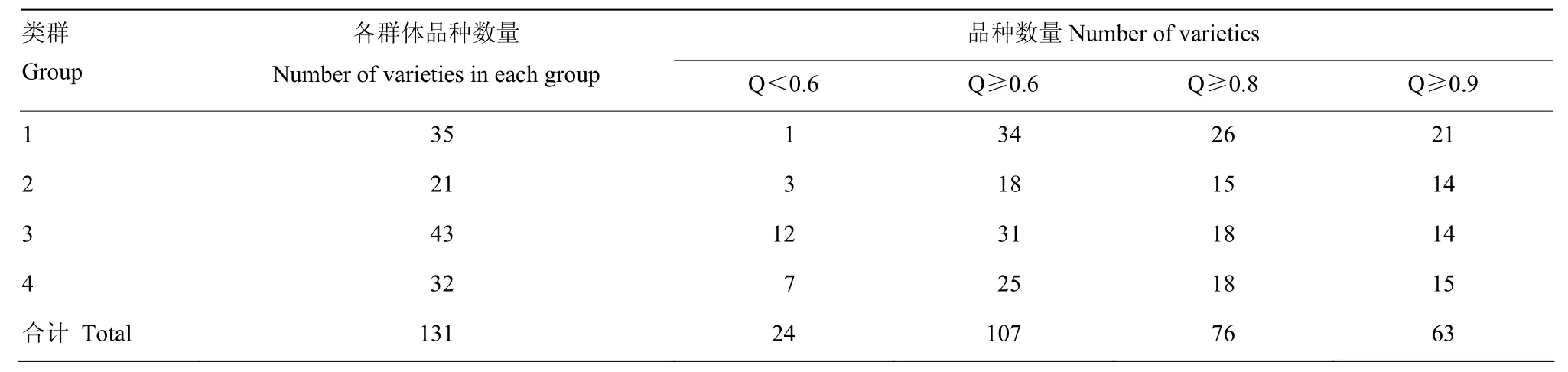
表5 各群体Q值分布Table 5 Distribution of Q-value of four groups
3 讨论
3.1 荧光SSR标记分析技术在糜子上的应用
SSR标记的检测可采用琼脂糖凝胶电泳、非变性聚丙烯酰胺凝胶电泳、变性聚丙烯酰胺凝胶电泳和毛细管电泳荧光检测法,因方法不同,结果也会有所差别。基于全自动基因分析仪的SSR荧光标记技术是用6-FAM、HEX、ROX和TAMRA 4种荧光染料中的一种对引物进行标记,扩增的PCR产物和标准分子量样品(内标)在同一毛细管泳道中电泳,利用软件进行图像收集和扩增片段的大小分析,实现了SSR标记与高效、自动化技术的结合,比传统聚丙烯酰胺凝胶电泳检测技术准确性更高,重复性更好。本试验同时采用变性聚丙烯酰胺和毛细管电泳双向检测技术分析22对引物在131份糜子材料中的遗传多样性,检测结果基本一致。但传统的变性聚丙烯酰胺凝胶电泳检测技术在不同板之间数据整合比较困难,当片段差异较小时,读带也有一定的困难,荧光SSR分子标记技术有效解决了不同板,不同实验室数据整合问题。
本研究所选用的22对标记中,有11对与连帅等[15]相同,这11对检测出的等位变异数平均为6.5个,而连帅用非变性聚丙烯酰胺凝胶电泳检测出的等位变异数平均为3个,这说明SSR荧光标记检测技术能检测到传统聚丙烯酰胺凝胶电泳发现不了的等位变异,结果更加准确可靠。
3.2 SSR标记在糜子种质资源遗传多样性研究中的应用
CHO等[13]基于糜子基因组DNA富集SSR文库开发了首批糜子的SSR标记,利用25个标记在50份材料中检测到110个等位变异,平均每个标记的等位变异数为4.4个,平均PIC为0.33。HUNT等[14]用其中的16个标记检测了98份欧亚大陆糜子地方品种的遗传多样性,平均每个位点检测到4.9个等位变异,平均基因多样性指数和多态性信息含量分别为 0.391和0.360。连帅等[15]用63对引物分析了来自国内外的192份糜子品种的遗传多样性,平均每个SSR位点检测到2.56个等位变异,平均PIC值为0.4855。王瑞云等[24]利用15个糜子特异性SSR标记检测来自中国11个省(区)的132份糜子材料,平均基因多样性指数和多态性信息含量分别为0.5298和0.4864。本研究选用22对SSR引物对131份糜子材料进行分析,共检测出128个主要等位变异,平均每个5.82个,基因多样性指数为 0.3572—0.8132,平均 0.6284;多态性信息含量为0.2934—0.8150,平均0.5874;Shannon多样性指数为0.5427—1.7681,平均1.2062。本试验所用SSR标记的多态性均高于前人研究结果。一方面本试验的引物经过两次筛选,引物多态性高,同时参试材料数量多、来源广,遗传多样性丰富;另一方面所用的检测方法分辨率更高,准确性更好。这也表明对已开发的糜子SSR标记进行进一步筛选的必要性,筛选高多态性引物用于糜子遗传多样性研究和DNA指纹图谱的构建。此外,建立高分辨率且成本较低的分子标记检测方法对于促进糜子遗传多样性研究也具有非常重要的意义。
3.3 中国糜子种质资源的遗传差异
中国糜子资源分布广泛,主要分布在七个生态区,分别为东北春糜子区、华北夏糜子区、北方春糜子区、黄土高原春夏糜子区、西北春夏糜子区、青藏高原春糜子区和南方秋冬糜子区。不同生态区,不同材料间的遗传多样性以及遗传关系成为近年来糜子研究的热点。王瑞云等[31]利用高基元微卫星标记分析了 96份糜子材料的遗传多样性并提出黄土高原和北方春糜子区的遗传多样性最丰富。连帅等[15]采用 63对高多态性SSR标记研究了来自于国内外的192份糜子地方品种和野生材料的遗传多样性,并发现内蒙古、东北、黄土高原地区种质资源遗传关系较其他地区更为复杂。薛延桃等[17]对近几年新收集引进的国外糜子地方品种和野生材料共计146份进行分析发现国内野生材料的遗传多样性高于国外地方品种,且河北群体的遗传多样性最为丰富。HU等[2]发现黄土高原地区的遗传多样性最丰富,可能是糜子的起源中心。董俊丽等[32]对糜子骨干种质进行遗传多样性分析发现遗传多样性与生态环境密切相关,且山西糜子资源的遗传多样性最丰富。本研究分析了来自5个生态区的131份糜子育成品种和农家种的遗传多样性和群体结构,发现北方春糜子区和黄土高原春夏糜子区的遗传多样性最丰富,与王瑞云等[31]结论一致。通过比较本研究与前人研究结果发现不同结果之间遗传差异存在较大差别,究其原因,可能有以下几个方面:一是各研究所采用的试验材料种类有所差异,如连帅等[15]和薛延桃等[17]文章中均包含了一定数量的国外品种以及国内野生资源,从而造成不同区域遗传多样性有所差异;二是不同研究中对同一生态区的试验材料选择数量的差别也会造成结果出现差异,如本研究选用北方春糜子区和黄土高原春夏糜子区的材料较多,西北春夏糜子区和南方秋冬糜子区的材料较少,为更加准确评估中国糜子资源的遗传多样性,选材需更加丰富。
群体结构分析结果表明,甘肃省材料 66.7%集中在群组Ⅰ中,青海省材料 87.5%聚在群组Ⅰ中,黑龙江省材料 66.7%聚在群组Ⅱ中,山西省的材料在各个组群中均有出现,与聚类结果基本一致。说明大部分糜子品种的遗传差异与地理来源相关,有些品种没有明显的区域特征,可能是不同区域间品种经过多次基因重组所致。
3.4 中国糜子育成品种和农家种的遗传多样性比较
国内外对糜子的遗传多样性和群体结构分析有很多[13-18],但有关中国糜子育成品种和农家种遗传差异的相关报道并不多见,本研究分析比较了来自中国不同生态区的 72份糜子育成品种及当地主要农家种的遗传多样性和群体遗传结构,发现北方春糜子区农家种的遗传多样性高于育成品种,东北春糜子区和黄土高原春夏糜子区农家种的遗传多样性略低于育成品种。聚类分析和群体结构分析结果表明,东北春糜子区的育成品种和农家种并不独立成群,在各组群中的分布基本一致,这可能因为糜子育成品种主要以农家种为遗传背景选育而来,而黄土高原春夏糜子区的育成品种和农家种分布在不同的组群中,这说明黄土高原春夏糜子区在育种过程中引种资源广泛,有效利用优良种质用于育种创新。北方春糜子区的农家种具有更高的遗传多样性,可为糜子新品种选育过程中挖掘优良基因和拓宽遗传基础提供重要的利用价值。
4 结论
北方春糜子区和黄土高原春夏糜子区的遗传多样性高于东北春糜子区,糜子的遗传差异与地理来源相关,东北春糜子区的育成品种主要以农家种为遗传背景选育而来,黄土高原春夏糜子区在育种过程中引种资源广泛,与其他生态区存在基因交流。
